

Journal of Environmental and Agricultural Sciences (JEAS). Ahmad et al., 2015. Volume 3: 14-22
Open Access – Research Article
Priming with Ascorbic Acid, Salicylic Acid and Hydrogen Peroxide Improves Seedling Growth of Spring Maize at Suboptimal Temperature
Ijaz Ahmad 1, Shahzad Maqsood Ahmad Basra 2, Safdar Hussain 3,*Syed Azhar Hussain 4,
Hafez-ur-Rehman 2, Abdul Rehman 5, Amjad Ali 5
1 Soil Conservation Officer, Agriculture Complex, Jail Road, Gujrat, 50800, Pakistan
2Department of Crop Physiology, University of Agriculture, 38040, Faisalabad, Pakistan
3 Department of Agronomy, Ghazi University, Dera Ghazi Khan, 32200, Pakistan
4 Agricultural Officer, Soil and Water Testing Laboratory, Sargodha, Pakistan
5 College of Agriculture, University of Sargodha, Sargodha, Pakistan
Abstract: Early sowing of maize crop can contribute to increase maize yield but poor stand establishment at low temperature is main hindrance in its productivity. Seed priming with ascorbic acid, salicylic acid and hydrogen peroxide improves seedling establishment at suboptimal temperature. Maize seeds were soaked in 20 and 40 mg L-1 aerated solution of ascorbic acid, salicylic acid and hydrogen peroxide for 24 hour and were dried back to its original weight. Primed and non-primed seeds were sown in pots containing sand under net house conditions. At low temperature, seed priming with hydrogen peroxide, ascorbic acid and salicylic acid improved seedlings’ growth probably through inducing superoxide dismutase (SOD) activities, better chlorophyll contents and enhanced nutrient contents. Seed priming with ascorbic acid, salicylic acid and hydrogen peroxide improved seedling establishment by inducing the antioxidants defense system and nutrient homeostasis. Seed priming with either 20 mg L-1 or 40 mg L-1 solution of H2O2, AsA and SA showed maximum seed invigoration and better performance in maize through inducing SOD activity and improving nutrient contents in root and shoot. The 20 mg L-1 of H2O2, salicylic acid and ascorbic acid seem to be suitable concentration for seed priming.
Keywords: Antioxidant activity, Chlorophyll, Nutrient contents, Plant growth, Seed priming, Temperature stress
*Corresponding authors: Safdar Hussain: safdarb1184@yahoo.com
Cite this article as Title: Priming with Ascorbic Acid, Salicylic Acid and Hydrogen Peroxide improves Seedling Growth of Spring Maize at Suboptimal Temperature Authors: Ijaz Ahmad, Shahzad Maqsood Ahmad Basra, Safdar Hussain, Syed Azhar Hussain, Hafez-ur-Rehman, Abdul Rehman and Amjad Ali Pages: 14-22
Ahmad, I., S.M.A. Basra, S. Hussain, S.A. Hussain, Hafeez-ur-Rehman, A. Rehman and A. Ali. 2015. Priming with ascorbic acid, salicylic acid and hydrogen peroxide improves seedling growth of spring maize at suboptimal temperature. Journal of Environmental & Agricultural Sciences. 3:14-22.[Abstract] [Citations]
Copyright © Ahmad et al., 2015. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium provided the original author and source are appropriately cited and credited.
Similar Articles Published in JEAS
- Zaib et al., 2022. Water Stress in Crop Plants, Implications for Sustainable Agriculture: Current and Future Prospects. Journal of Environmental & Agricultural Sciences. 24: xx-xx. [Abstract] [View Full-Text]
- Zahalan, R. and M.M. Alzoubi. 2021. Effect of Organic Physical Soil Amending on Deficit Irrigation Efficiency of potato (Solanum Tuberosum L.). Journal of Environmental & Agricultural Sciences. 23(1&2): 11-18. [ View Full-Text ] [Citations]
- Nasir, M.W., A. Yasmeen, M. Imran and T. Zoltan. 2019. Seed priming to alleviate drought stress in cotton. Journal of Environmental& Agricultural Sciences. 21:14-22.
[Abstract] [View Full-Text] [Citations] - Mazhar, M.S., B.E. Bajwa, G. McEvilly, G. Palaniappan and M.R. Kazmi. 2019. Improving vegetable value chains in Pakistan for sustainable livelihood of farming communities. Journal of Environmental & Agricultural Sciences. 18:1-9.. [Abstract] [View Full-Text] [Citations]
- Ahmad, I., S.M.A. Basra, S. Hussain, S.A. Hussain, Hafeez-ur-Rehman, A. Rehman and A. Ali. 2015. Priming with ascorbic acid, salicylic acid and hydrogen peroxide improves seedling growth of spring maize at suboptimal temperature. Journal of Environmental & Agricultural Sciences. 3:14-22. [ ] [View Full-Text] [Citations]
1. Introduction
Maize is chilling sensitive crop during early growth stage. Chilling stress is a key environmental factor which limits growth of crop plants in response to the over production of reactive oxygen species (ROS). ROS are known to exacerbate imbalance between light absorption and its utilization by inhibiting Calvin-Benson cycle activity (Logan et al., 2006). ROS also reduce content and activity of ribulose bisphosphate carboxylase oxygenase (RUBISCO) enzyme which leads to higher electron flux to O2 coupled with reduced CO2 accumulation (Zhou et al., 2006). Chilling-induced oxidative stress in maize can lead to increased accumulation of ROS which ultimately damage the macromolecules like lipid and protein (Sokolnik et al., 2009; Prasad, 1997). In addition, responses to chilling-induced oxidative stress are alteration in activities of enzymes of antioxidant defense system. However, if the duration of chilling stress is too long, the defense system may not remove overproduced ROS effectively, which may result in severe oxidative damage or even death of cells (Sokolnik et al., 2009). Several endogenous defense mechanisms both enzymatic and non-enzymatic are involved in scavenging ROS (Rayduk et al., 2009). Enzymatic system consists of a number of enzymes such as superoxide dismutase, ascorbate peroxidase and catalase and in nonenzymic mechanism; metabolites like ascorbic acid, salicylic acid and low concentration of H2O2 are of prime importance that scavenge these radicals and protect membranes from injurious effects of ROS (Sharma et al., 2012; Ahmad et al., 2012). Many comparative studies using chilling tolerant and sensitive genotypes have shown that greater antioxidant capacity was exhibited in chilling-tolerant species compared to sensitive ones (Jahnke et al., 2009).
Seed priming is an important tool to improve (Batool et al., 2015) crop vigor. Seed Priming has been used to improve the crop establishment in many crops at suboptimal temperature (Yoon et al., 1997; Dashtamian et al., 2014). Seed priming with salicylic acid improved chilling tolerance by increased germination (Sedghi et al., 2010), activation of antioxidants, maintenance of tissue water contents and reduced membrane permeability (Farooq et al., 2008). Seed priming with ascorbic acid, salicylic acid and H2O2 induced salinity tolerance (Athar et al., 2008; Gautam and Singh, 2009; Wahid et al., 2007; Kumar et al., 2010; Ahmad et al., 2012). Thus, it seems that ascorbic acid, salicylic acid and hydrogen peroxide are promising materials for seed treatments.
In the view of previous work done on horticultural crops like pansy and pepper, it can be assumed that seed priming can provide early establishment of vigorous and healthy maize seedlings at suboptimal temperature. To the best of our knowledge, limited work has been reported regarding biochemical and physiological changes in maize induced by seed priming. Therefore, the present study was carried out to investigate the effect of seed priming with different concentration of ascorbic acid, salicylic acid and hydrogen peroxide on enhancing germination and seedling vigor of maize hybrid and to explore the biochemical basis of this enhancement.
2. Material and Methods
The experiment was carried out in the wire house of Department of Crop Physiology, University of Agriculture, Faisalabad, Pakistan by employing completely randomized design (CRD) with three replications. Healthy and uniform seeds of maize hybrid (HI GEN Sawn 9697) were selected for experimental study.
2.1 Seed Priming
The solution of 20 and 40 mg L-1 of ascorbic acid (AsA), salicylic acid (SA) and hydrogen peroxide (H2O2) were prepared in distilled water. For priming, maize seeds were soaked in aerated solution of respective osmoticum for 24 h at room temperature. Seed weight to solution volume ratio was 1:5 (w/v). During priming fresh air was supplied with aeration pump continuously. After each treatment, seeds were rinsed thoroughly with distilled water, spread on thin layer of filter paper and dried back closer to original weight. These seeds were packed in polythene bags and stored in a refrigerator at 5°C until use.
2.2 Seed Sowing
Primed seeds as per treatment were sown in pots containing sand while untreated seeds were taken as control. Hoagland solution was used to nourish the plants uniformly throughout course of study. The atmospheric temperature was recorded from sowing to harvesting and weekly average were computed and presented (Table 1). At 4th leaf stage plants were harvested and tested for seedling vigor, antioxidants and nutrient analysis.
Table 1: Weekly average atmospheric temperature recorded during course of studies

2.3 Seedling growth and vigor
Five seedlings were carefully harvested from each pot, shoot and root length of every plant was measured with measuring tape and their averages were recorded for statistical analysis. Root length obtained in above parameter was divided by the shoot length to get the root: shoot ratio.
2.4 Determination of Superoxide Dismutase
To extract antioxidant enzymes, 0.5 g fresh leaves randomly sampled from plants in each pot were harvested and immediately ground using a tissue grinder in 8 mL of cooled phosphate buffer [pH 7.0, containing 1% (w/v) polyvinylpyrrolidone] and 0.2 g quartz sand in test tubes that were placed in an ice bath. The homogenate was centrifuged at 15000-x g for 20 minutes at 4oC. The supernatant was used for assays of enzyme activity. The activity of SOD was determined by measuring its ability to inhibit the photoreduction of nitro blue tetrazolium (NBT) following the method of Giannopolitis and Ries (1977). The reaction solution (3 mL) contained 50 uM NBT, 1.3 uM riboflavin, 13 mM methionine, 75 nM EDTA, 50 mM phosphate buffer (pH 7.8) and 20 to 50 uL enzyme extract. Test tubes containing the reaction solution and leave were irradiated under light bank (15 flouresent lamps) at 78 u mol m-2 s-1 for 15 minutes. The absorbance of irradiated and non-irradiated solution at 560 nm was determined with spectrophotometer (T60 spectrophotometer). One unit of SOD activity was defined as the amount of enzyme that would inhibit 50 % of NBT photo reduction.
2.5 Determination of Chlorophyll Contents
The fresh leaves were cut into 0.5 cm segments and extracted overnight with 80 % acetone at -10oC. The extract was centrifuged at 14000 g for 5 min and the absorbance of supernatant was recorded at 645 and 663 nm using a spectrophotometer (T60 U spectrophotometer PG Instruments, Limited). The chlorophyll a (Chl a) and b (Chl b) contents were calculated by using formulae given by Nagata and Yamashita (1992).
2.6 Determination of Nitrogen Contents
Total nitrogen was estimated by Kjeldhal apparatus. 10 mL of digested sample was taken in Kjeldhal flask and placed it on the Micro-Kjeldhal ammonium distillation unit and then 10 mL of 40% sodium hydroxide solution were added and immediately flask was connected to distillation apparatus. 10 mL 4% boric acid was taken along with mixed indicator in 100 mL conical flask. When distillate was approximately 40-50 mL, conical flask was removed and distillation was turned off. The distillate was cooled for a few minutes and titrated against 0.01N H2SO4 up to end point. Nitrogen was calculated by the formula given by Singh et al. (2005).
2.7 Determination of Phosphorus Contents
Phosphorus was determined by using a spectrophotometer. The digestion material (2 ml) was dissolved in 2 ml of Barton reagent and maintains volume up to 50 ml. The samples were then subjected to spectrophotometer (Spectrophotometer AnA-720 W Japan) and recorded the absorbance at 470 nm for P ions. Run a blank (without P) simultaneously. Prepared standard solution of KH2PO4 (Potassium Di-Hydrophosphate) and formed the standard curve by plotting P-concentration on X-axis and percent transmission/ spectrometer readings on Y-axis. P was calculated by following formula produced by Singh et al. (2005).
2.8 Determination of Potassium Contents
Potassium concentration was determined by using Flame photometer-410 (Corning Model). A graded series of standards (10 to 40 mg L-1 of K) were prepared and standard curve for standard was plotted by K-concentration on X-axis and flame photometer on Y-axis. The O.D values of K+ from flame photometer were compared with standard curve and concentration of K+ element was computed (Singh et al., 2005).
2.9 Statistical Analysis
The data collected was analyzed statistically using Fisher’s analysis of variances technique and treatment means showing F-values significant compared using least significance difference at 0.05 probability level (Steel et al., 1997).
Figures sharing same letter, in each column, did not differ significantly at 0.05 level of probability; Chl a = Chlorophyll a content; Chl b = Chlorophyll b content; SOD = Superoxide dismutase; AsA = Ascorbic acid; SA = Salicylic acid and H2O2 = Hydrogen peroxide. AsA = Ascorbic acid; SA = Salicylic acid and H2O2 = Hydrogen peroxide.
3. Results
3.1 Seedling vigor and biochemical analysis
Seed priming with different levels (20 and 40 mg L-1) of ascorbic acid, salicylic acid and hydrogen peroxide, significantly affected root and shoot length. Seedlings dry weight and length were improved by different priming strategies but seed primed with AsA-40 H2O2-40 were leading in improving dry weight and seedling length, respectively. It is evident from comparison means that all priming strategies improved root and shoot length (Table 2). This increase in root length and shoot length were in order of H2O2-20> H2O2-40> AsA-40> SA-40> AsA-20 and H2O2-40> AsA-40> AsA-20> H2O2-20> SA-40> SA-20, respectively. The comparison of different priming techniques indicates that all priming strategies improved root length in maize except SA-20 with similar root length (Table 2).
Table 2 Seedling dry weight, seedling length, root length, shoot length and root shoot ratio Chl a, Chl b and SOD activity of maize as influenced by different seed priming treatments

Root shoot ratio was reduced significantly by difference priming treatments. The lowest root shoot ratio in seedling was produced when seeds were treated with ascorbic acid and salicylic acid (Table 2).
3.2 Chlorophyll contents (mg 100mL-1)
Plant photosynthetic efficiency depends on pigments like Chl a and Chl b involved in photosynthesis. All priming strategies improved leaf Chl a and b contents (Table 2). Maximum Chl a content (2.68 mg 100mL-1) were observed from seeds primed with SA-20 which was statistically at par with pre-soaked seeds with AsA-20, SA-40, H2O2-40, AsA-40 and H2O2-20. However, minimum Chl a contents (2.35 mg 100mL-1) were noted for control. Data depicts that all the priming treatments improved Chl b contents (Table 2) except non-primed control with minimum Chl b contents of 0.93. Nonetheless, there was no difference among priming treatments for this trait.
3.3 Superoxide dismutase (unit mg-1 Protein)
Superoxide dismutase (SOD) is the most effective enzyme in preventing cellular damage by converting superoxide radical to H2O2.In present study, chilling stress caused reduction in SOD activity of maize but the seed priming with ascorbic acid, salicylic acid and hydrogen peroxide enhanced SOD activity. Higher SOD activity in leaves under chilling stress suggests a more efficient scavenging system which may result in better protection against ROS during stress (Table 2).

Figure 1. Root nitrogen (a), phosphorous (b) and potassium (c) contents in response to different priming strategies
3.4 Nutrient analysis
Root and shoot nitrogen (N), phosphorus (P) and potassium (K) contents were significantly reduces by chilling stress. However, seed primed with ascorbic acid, salicylic acid and hydrogen peroxide significantly improved the absorption of N, P and K in root (Fig. 1a, b &c) and uptake in shoot (Fig. 2a, b& c). Different priming strategies significantly increased N, P and K contents in root and shoot, but different priming techniques showed similar results in improving nutrient contents.
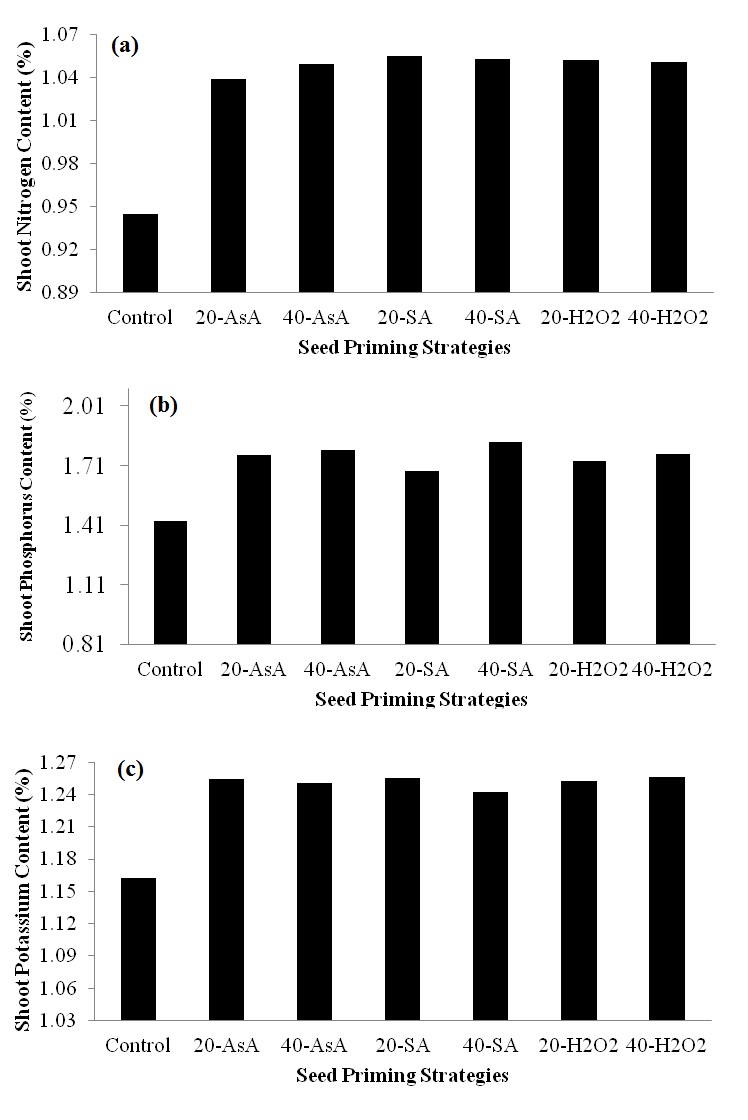
Figure 2. Shoot nitrogen (a), phosphorous (b) and potassium (c) contents in response to different priming strategies
4. Discussion
4.1 Seedling vigor and biochemical analysis
Chilling stress is a key environmental factor which limits growth of crop plants in response to the over production of ROS by inhibiting activity of Calvin-Benson cycle (Logan et al., 2006), enhancing over reduction of respiratory electron transport chain (Hu et al., 2008) and by reducing activity of RUBISCO (Zhou et al., 2006). Seedling growth in term of both weight and length was increased in seed primed treatment with ascorbic acid, salicylic acid and hydrogen peroxide possibly by ameliorating injurious effects of ROS on light harvesting apparatus in maize plants subjected to chilling stress (Table 2). This view is supported by a positive relationship between seedling dry weight and Chl a and SOD activity (Fig. 4 and 5). H2O2 at low concentration in biological systems acts as a signal molecule in cells (Neill et al., 2002; Kumar et al., 2010), ascorbic acid (AsA) is the most important antioxidants (Shalata and Neumann, 2001). Salicylic acid (SA) regulates cell growth by expansion and protecting the cell structure (Kang et al., 2007).
Low temperature lowers cell elongation (Ben-Haj-Salah and Tardieu, 1995) but in the present study shoot and root length increased significantly by different priming strategies and increased in shoot and root growth might be due to higher cell expansion in primed seeds as compared to unprimed seeds (Table 2). Similar findings were described by Kaydan et al, (2007) who reported that primed seeds with SA enhance the root and shoot growth under salinity stress.
Priming strategies improved leaf Chl a and b contents (Table 2). Low temperature (<10oC) induce chilling injury through production of reactive oxygen species (ROS) which causes oxidative damage to chlorophyll molecule (Guan et al., 2009). This was confirmed by increased SOD activities in seeds that were primed with SA, AsA and H2O2 (Table 2). These results suggested that seed priming with AsA, SA and H2O2 could be used to minimize the harmful effect of low temperature on leaf chlorophyll contents which consequently may enable the plant to withstand at suboptimal conditions.

Figure 3. Relationship between Chlorophyll a and seedling dry weight

Figure 4. Relationship between superoxide dismutase and seedling dry weight
Responses to chilling-induced oxidative stress include alteration in activities of enzymes of antioxidant defense system. However, chilling stress caused reduction in SOD activity of maize but the seed priming with ascorbic acid, salicylic acid and hydrogen peroxide enhanced SOD activity (Table 2). Higher SOD activity in leaves under chilling stress suggests a more efficient scavenging system which may result in better protection against ROS during cold stress. Many studies indicated that chilling-tolerant genotypes have greater antioxidant capacity than sensitive ones (Jahnke et al., 2009; Hodges et al., 1996; Huang; Chen and Arora, 2012; Guo, 2005). Seed priming with AsA (Athar et al., 2008), SA (Gautam and Singh, 2009) and H2O2 (Kumar et al., 2010; Wahid et al., 2007a) also proved that these priming techniques induce SOD activity which quench oxygen radicals under stress (Foyer and Noctor, 2003: Orabi et al., 2015).
Chilling-induced oxidative stress evident by increased accumulation of ROS, including H2O2 and O2•−, lipid peroxidation, and protein oxidation is a significant factor in relation to chilling injury in plants (Fryer et al., 1996; Prasad, 1997; Sokolnik et al., 2009). Protein carbonyl content, an indication of oxidative damage, was increased 2-fold in maize seedlings when exposed to chilling temperatures (Prasad, 1997). Lipoxygenase activity as well as lipid peroxidation was increased in maize leaves during low temperatures, suggesting that lipoxygenase-mediated peroxidation of membrane lipids contributes to the oxidative damage occurring in chill stressed maize leaves (Fryer et al., 1996).
Low temperature limits root growth, which reduce water and nutrient absorption (Stamp et al. 1997). Seed priming treatments with AsA, SA and H2O2 significantly improved the root N, P and K contents significantly (Fig.1a, b and c) by reducing the deteriorative oxidative injury caused by ROS in root system. Results of present experiment clearly indicated that primed seeds effectively improved nutrient absorption and root growth, possibly by enhancing SOD activity. These results are consistent with those Gunes et al. (2007) and Wahid et al. (2007) who reported that seed primed with SA increased N and K contents in maize while seed primed with H2O2increased P contents in wheat under saline conditions, respectively.
Low temperature reduced root length (Kaspar and Bland, 1992) and shorter roots limited the phosphorous uptake efficiency (Enns et al. 2006) but pre-soaking seed treatments with ascorbic acid, salicylic acid and hydrogen peroxide increased shoot N, P and K content at suboptimal growth temperature (Fig. 2a, b and c). Improved shoot N, P and K contents may be explained by improved and well developed root system, which resulted in improved nutrients uptake and its translocation towards shoot parts. These results also support the findings of Wahid et al. (2007) and Sakr and Arafa (2009) who reported that seed primed with H2O2 and SA reduced NO3–,P and K+ in wheat and canola under stress conditions (Gunes et al., 2007; Wahid et al., 2007; Sakr and Arafa , 2009). These results also support the findings of Wahid et al. (2007), and Sakr and Arafa (2009) who reported seed primed with H2O2 and SA reduced membrane permeability and leakage of ions like NO3– in wheat and canola under saline conditions. In the present study the increased accumulation of potassium (K+) in maize seedlings seems to be responsible for their survival under chilling stress playing important role in osmotic adjustment.
5. Conclusion
On the basis of results obtained from this investigation it may be concluded maize responded similarly to different seed priming treatments. Seed priming with either 20 mg L-1 or 40 mg L-1 solution of H2O2, AsA and SA showed maximum seed invigoration and better performance in maize through inducing SOD activity and improving nutrient contents in root and shoot. The 20 mg L-1 of H2O2, salicylic acid and ascorbic acid seem to be suitable concentration for seed priming.
Acknowledgments: We acknowledge the co-operation of the staff working at Crop Physiology Department Laboratory, University of Agriculture, Faisalabad for helping in plant samples analysis.
Competing Interests: Authors declare that they have no competing interests.
References
Ahmad, I., T. Khaliq, A. Ahmad, S. M. A. Basra, Z. Hasnain and A. Ali. 2012. Effect of seed priming with ascorbic acid, salicylic acid and hydrogen peroxide on emergence, vigor and antioxidant activities of maize. Afr. J. Biotechnol.11: 1127-1137.
Apel, K. and H. Hirt. 2004. Reactive oxygen species: metabolism, oxidative stress and signal transductions. Annual Rev. Plant Biol. 55: 373-399.
Athar, H. R., A. Khan and M. Ashraf. 2008. Exogenously applied ascorbic acid alleviates salt induced oxidative stress in wheat. Environ. Exp. Bot. 63: 224-231.
Batool, A., K. Ziaf and M. Amjad. 2015. Effect of halo-priming on germination and vigor index of cabbage. J. Environ. Agri. Sci. 2:8.
Ben-Haj-Salah, H. and F. Tardieu. 1995. Temperature affects expansion rate of maize leaves without change in distribution of cell length. Plant Physiol. 109: 861-870.
Chen, K. and R. Arora. 2013. Priming memory invokes seed stress tolerance. Environ. Exp. Bot. 94:33-45.
Dashtamian, P. N., M. K. Hasseini and M. Esfahani. 2014. Improving of chilling tolerance of rice seedling by seed priming with salicylic acid. Archi. Agron. Soil Sci. 60: 1291-1302.
Enns, L. C., M. E. McCully and M. J. Canny. 2006. Branch roots of young maize seedlings, their production, growth and phloem supply from the primary root. Func. Plant Biol. 33: 391-399.
Farooq, M., T. Aziz, S. M. A. Basra, M. A. Cheema and H. Rehman. 2008. Chilling tolerance in hybrid maize induced by seed priming with salicylic acid. J. Agron. Crop Sci. 194:161-168.
Foyer, C.H., and G. Noctor. 2003. Redox sensing and signaling associated with reactive oxygen in chloroplast, peroxisomes and mitochondria. Physiol. Plant. 119: 355-364.
Fryer, M. J., J. R. Andrews, K. Oxborough, D. A. Blowers and N. R. Baker. 1998. Relationship between CO2 assimilation, photosynthetic electron transport, and active O2 metabolism in leaves of maize in the field during periods of low temperature. Plant Physiol. 116: 571-580.
Gautam, S. and P. K. Singh. 2009. Salicylic acid induced salinity tolerance in corn under NaCl stress. Acta Physiol. Plant. 31: 1185-1190.
Giannopolitis, C. N. and S. K. Ries. 1977. Superoxide dismutase. I. Occurrence in higher plants. Plant Physiol., 59: 309-314.
Guan, Y. J., J. Hu, X. J. Wang and C. X. Shao. 2009. Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress. J. Zhejiang Univ. Sci. B. 10: 427-433.
Gunes, A., A. Inal, M. Alpaslan, F. Eraslan, E.G. Bagci and N. Cicek. 2007. Salicylic acid induced changes on some physiological parameters symptomatic for oxidative stress and mineral nutrition in maize (Zea mays L.) grown under salinity. J. Plant Physiol., 164: 728-736.
Hodges, D. M., C. J. Andrews, D. A. Johnson and R. I. Hamilton. 1996. Antioxidant compound responses to chilling stress in differentially sensitive inbred maize lines. Physiol. Plant. 98: 685-692.
Hu, W. H., X. S. Song, K. Shi, X. J. Xia, Y. H. Zhou and J. Q. Yu. 2008. Changes in electron transport, superoxide dismutase and ascorbate peroxidase isoenzymes in chloroplasts and mitochondria of cucumber leaves as influenced by chilling. Photosynthetica. 46: 581-588.
Huang, M. and Z. Guo. 2005. Responses of antioxidative system to chilling stress in two rice cultivars differing in sensitivity. Biol. Plant. 49: 81-84.
Jahnke, L. S., M. R. Hull and S. P. Long. 1991. Chilling stress and oxygen metabolizing enzymes in Zea mays and Zea diploperennis. Plant Cell Environ. 14: 97-104.
Kang, G. Z., Z. X. Wang, K. F. Xia and G. C. Sun. 2007. Protection of ultrastructure in chilling-stressed banana leaves by salicylic acid. J. Zhejiang Univ. Sci. B. 8: 277-282.
Kaspar, T. C. and W. L. Bland. 1992. Soil temperature and root growth. Soil Sci. 154: 290-299.
Kaydan, D., M. Yağmur, N. Okut and G. Tarihi. 2007. Effects of salicylic acid on the growth and some physiological characters in salt stressed wheat (Triticum aestivum L.). Tarim Bilimleri Dergisi. 13: 114-119.
Kumar, M., G. Sirhindi, R. Bhardwaj, S. Kumar and G. Jain. 2010. Effect of exogenous H2O2 on antioxidant enzymes Brassica juncea L. seedlings in relation to 24-epibrassinolide under chilling stress. Ind. J. Biochem. Biophysics. 47: 378-382.
Logan, B.A., D. Kornyeyev, J. Hardison and A. S. Holaday. 2006. The role of antioxidant enzymes in photoprotection. Photosynthesis Res. 88: 119-132.
Orabi, S. A., M. G. Dawood and S. R. Saleem. 2015. Comparative study between the physiological role of hydrogen peroxide and salicylic acid in alleviating low temperature stress on tomato plants grown under sand-ponic culture. Sci. Agriculturae. 9(1): 49-59.
Nagata, M. and I. Yamashita.1992. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. J. Japanese Soc. Food Sci. Technol. 39: 928-928. (In Japanese with English abstract).
Neill, S.J., R. Desikan, A. Clarke, H. D. Hurst and J. T. Hancock. 2002. Hydrogen peroxide and nitric oxide as signaling molecules in plants. J. Exp. Bot. 53: 1237-1247.
Prasad, T. K. 1997. Role of catalase in inducing chilling tolerance in pre-emergent maize seedlings. Plant Physiol. 114: 1369-1376.
Radyuk, M. S., I. N. Domanskaya, R. A. Shcherbakov and N. V. Shalygo. 2009. Effect of low above-zero temperature on the content of low-molecular antioxidants and activities of antioxidant enzymes in green barley leaves. Russian J. Plant Physiol. 56:175-180.
Sakr, M. T. and A. A. Arafa. 2009. Effect of some antioxidants on canola plants grown under soil salt stress condition. Pak. J. Biol. Sci. 12: 582-588.
Sedghi, M., A. Nemati and B. Esmailepour. 2010. Effect of seed priming on germination and seedling growth of two medicinal plants under salinity. Emir. J. Food Agric. 22: 130-139.
Shalata A. and P. M. Neumann. 2001. Exogenous ascorbic acid (Vitamin C) increases resistance to salt tolerance and reduced lipid peroxidation. J. Exp. Bot. 364: 2207-2211.
Sharma, P., A. B. Jha, R. S. Dubey and M. Pessarakli. 2012. Reactive oxygen species, oxidative damage and antioxidative defense mechanism in plants under stressful conditions. J. Bot. Article No. 217037. p: 1-26
Singh, D., P.K, Chhakarand B.S. Dwivedi. 2005. Manual on Soil plant and water analysis. Westville Publishing House, New Dehli, India. p. 77-92.
Sokolnik, A. Z., H. Asard, K. G. Koplinska and R. J. Gorecki. 2009. Cadmium and zinc-mediated oxidative burst in tobacco BY-2 cell suspension cultures. Acta Physiol. Plant. 31: 43-49.
Stamp, P., B. Feil, M. Schortemeyer and W. Richner. 1997. Responses of roots to low temperatures and nitrogen forms. In: Plant roots from cells to systems. (Ed.HM Anderson) Kluwer Academic Publishers: Dordrecht, The Netherlands. p. 143-154.
Steel, R. G. D., T. H. Torrie and D. A. Dickey. 1997. Principles and Procedures of Statistics. A Biometrical Approach 3rd Ed. McGraw Hill Book International Co. Inc., Singapore. p. 204-232.
Wahid, A., M. Perveen, S. Gelani and S. M. A. Basra. 2007. Pretreatment of seed with H2O2 improves salt tolerance of wheat seedlings by alleviation of oxidative damage and expression of stress proteins. J. Plant Physiol. 164: 283-294.
Yoon, B., H. J. Lang and B. G. Cobb. 1997. Priming with salt solutions improves germination of pansy seed at high temperatures. Hort. Sci. 32: 248-250.
Zhang, Y., Y. Luo. Y. X. Hou, H. Jiang, Q. Chen and R. H. Tang. 2008. Chilling acclimation induced changes in the distribution of H2O2 and antioxidant system of strawberry leaves. Agric. J. 3 286-291.
Zhang, Y., H. R. Tangand Y. Luo. 2008. Variation in antioxidant enzyme activities of two strawberry cultivars with short term low temperature stress. World J. Agric. Sci. 4: 458-462.
Zhou, Y. H., J. Q. Yu, W. H. Mao, L. F. Huang, X. S. Song and S. Nogues. 2006. Genotypic variation of Rubisco expression, photosynthetic electron flow and antioxidant metabolism in the chloroplasts of chill-exposed cucumber plants. Plant Cell Physiol. 47: 192-199.
Similar Articles Published in JEAS