
Journal of Environmental and Agricultural Sciences (JEAS). Rehman, 2020. Volume 22(3): 1-6
Open Access – Research Article
Abundance of Autumn Bees and their Natural Enemies in Sub-Tropical Bahawalpur, Pakistan
Aziz-Ur-Rehman 1, Asif Sajjad 1,*, Waseem Akram 1, Ammad Ahmad 1
1 Department of Entomology, University College of Agriculture and Environmental Sciences, The Islamia University, Bahawalpur, Pakistan
Abstract: Autumn season in Pakistan lasts from October to November. During these months most of the bees decline in their populations, and by the end of November, they start getting into hibernation to endure upcoming winter. Present study was carried out to investigate the abundance of different ground and cavity-nesting bees of the autumn season at the Baghdad-ul-Jadeed campus of the Islamia University of Bahawalpur, Punjab, Pakistan during 2019. The population of their natural enemies (predatory and parasitic wasps and cleptoparasitic cuckoo bees) and their floral host plants were also recorded. A total of ten bee species (including three cleptoparasitic cuckoo bees) and nine predatory or parasitic wasp species were recorded, however; bees were more abundant than wasps. Ground-nesting bees comprised 90.09% of total bee abundance. Wasps were higher in abundance than cleptoparasitic cuckoo bees. A cavity and a soil-nesting bee, i.e. Ceratina sp. and Pseudapis sp.1, respectively visited the maximum number of plant species. The maximum number of bee species visited Leucaena leucocephala, Trianthema portulacastrum and Carthamus oxyacantha. Current study gives a baseline for future conservation and pollination management programs in the region.
Keywords: native bees, ground-nesting, cavity-nesting, cuckoo bees, floral sources.
*Corresponding author: Asif Sajjad: asifbinsajjad@gmail.com
Cite this article as: Rehman, A.U., A. Sajjad, W. Akram and A. Ahmad. 2020. Abundance of autumn bees and their natural enemies in sub-tropical Bahawalpur, Pakistan. Journal of Environmental & Agricultural Sciences. 22(3): 1-6.

Copyright © Rehman et al., 2020. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium provided the original author and source are appropriately cited and credited.
1. Introduction
More than 20,000 species of the bees have been taxonomically described (Michener, 2000). Wild bees pollinate 87% of flowering plants and ensure 35% of the global food supply by pollinating 75% of global food crops (Ollerton et al., 2011). Not only in the wild and agricultural habitats, but they also accomplish pollination of plant species in urban habitats, i.e. public parks and re-vegetated wasteland (Cane, 2005). Several bee species have been documented as the effective pollinators of fruits and vegetables in southern Punjab, Pakistan (Akram et al. 2019; Zameer et al. 2017; Ali et al. 2014). Bees are usually hairier and perform different functions than wasps, such as the collection of pollen from flowers as a protein source (Michener, 2000). The diversity, abundance and richness of bees are reported to enhance the crop yield (Goulson, 2003). Previous studies have established a significant relationship between pollinators decline and decreased ecosystem service of crop pollination (Hausmann and Petermann, 2016).
Processes occurring at the landscape scale have a substantial influence on local diversity and dynamics (Kareiva and Wennergren, 1995; Polis et al., 1997). Habitat loss, fragmentation, agrochemicals, pathogens, alien species and climate change are mainly responsible for the decline in pollinator species (Potts et al., 2010). Organic cropping can be one of the conservation strategies of bees as it promotes the diversity of wild bees and density of managed bumble bees (Holzschuh et al., 2008). The reduction of bee population has led to serious concerns over ecosystem service of pollination and crop yield. Therefore, there is a need to conserve, monitor and maintain their population, especially in agricultural ecosystems (Perfecto and Vandermeer, 1996, Kremen et al., 2002).
As most of the native bees hibernate in the winter season, late autumn is a critical time to help conserve native bees by adopting some suitable conservation measure. However, this requires strong baseline knowledge of their populations, floral host plants, nesting requirements and natural enemies. Current study was planned in the same context aiming to document the diversity and assemblage of ground and cavity-nesting bees, their floral host plants and natural enemies during the autumn season (a time when most of the bees are preparing for hibernation) under the sub-tropical climatic conditions of Bahawalpur, Pakistan.
2. Materials and Methods
2.1. Study area
The study was conducted in Baghdad-ul-Jadeed campus of the Islamia University of Bahawalpur (IUB), Punjab, Pakistan (29°22’16.3″N 71°45’52.9″E; 181 meters above sea level), scattered over an area of 518 hectares. Climate of the study area is sub-tropical with a long hot summer and short cold winter. There are four different seasons: spring (March to May), summer (June to September), autumn (October to November) and winter (December to February), in the study area. Most of the plant species (about 60%) blossom during the spring season (Sajjad and Saeed, 2010; Sajjad et al., 2012). The campus is characterized by a variety of landscapes and land-use-types including naturally occurring desert, planted forest, agricultural land, ornamental grassy plots, roads and buildings.
2.2. Survey of bees
For observing soil-nesting bees, random walks were made in the study area by four observers from 10:00 h to 16:00 h on a weekly basis (i.e. two hours per observer per census). Total sampling effort was 72 hours around 12 observation days. During each census, each observer performed a random walk and observed any bee or wasp entering or leaving some hole in the soil. The bees or wasps were caught with a collection net and preserved in the laboratory of Department of Entomology, University College of Agricultuer and Environmental Sciences, Islamia University, Bahawalpur, for later taxonomic identification. Similar procedure and sampling efforts were made for cavity-nesting bees while observing dry wood logs and mud wall nesting in the study area.
2.3. Visitation census of bees on flowering plants
In order to know bee host plants, a weekly census was also conducted on all the available plants in the flowering phase. These include all trees, shrubs, seasonal weeds, ornamental plants, orchard trees and vegetables. In each census, five individuals of each plant species were randomly selected, and each plant was observed for five minutes for visitation of bees during peak activity timing (9:00 h to 11:00 h). The temperature and relative humidity during the whole study period (September to November) ranged from 22 °C to 40 °C and 51% to 74%, respectively. The visiting bees were collected, killed and preserved in the laboratory for later taxonomic identification.
2.4. Taxonomic Identification
Bees were identified to genus level by using keys of Michener (2007). Wasps were identified to family level by using keys of O’neill (2008).
3. Results
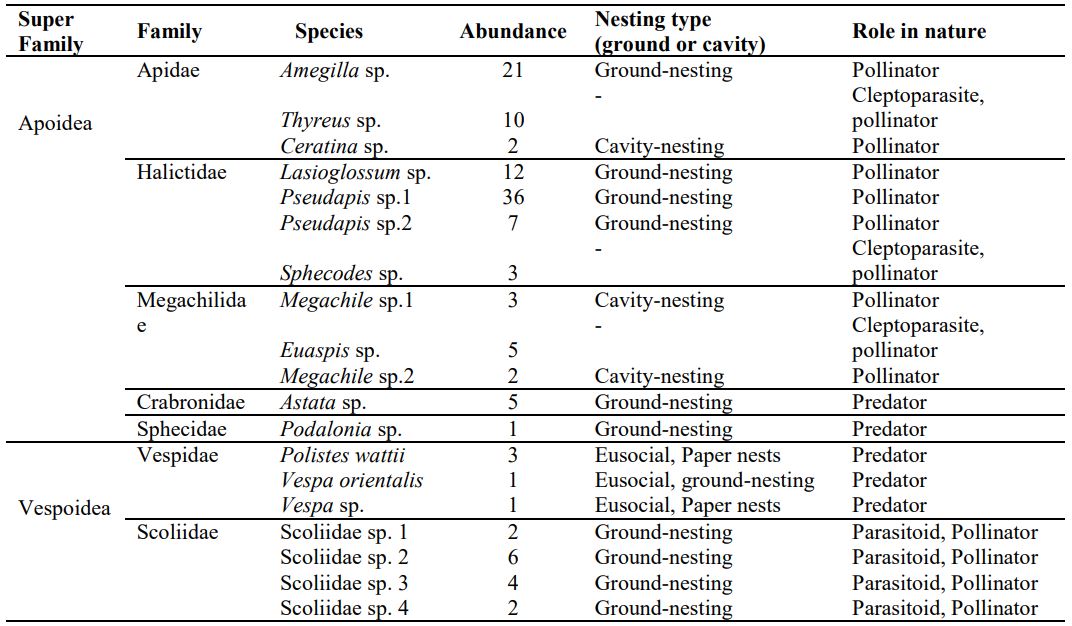
A total of ten bee species in three families (i.e. Megachilidae, Halictidae, Apidae) and nine wasp species in four families (i.e. Crabronidae, Scoliidae, Sphecidae, Vespidae) were recorded. A total of 125 individual were recorded, including 99 bees and 26 wasps. The soil-nesting bees were dominant over cavity-nesting bees in terms of their abundance, i.e. 90.09 % ground-nesting bees and 9.90% cavity-nesting bees. Two ground-nesting bees, i.e. Pseudapis sp.2 and Amegilla sp. comprised 57% of total bee abundance. Family Halictidae was the most abundant among all other families of bees.
Cleptoparasitic cuckoo bees (i.e. they do not make their nests; instead they lay their eggs in the nests of their host bees) belonged to genus Euaspis (Megachilidae), Thyreus (Apidae) and Sphecodes (Halictidae). The parasitic wasps (i.e. they usually lay their eggs in the larvae of their hosts) belonged to genus Podalonia (Sphecidae) while Predatory wasps belonged to genus Polistes, Vespa and Astata (Table 1). The ratio between pollinating bees and cleptoparasitic cuckoo bees was 8.8:1.3 and the ratio between pollinating bees and predatory wasps was 8.8:1. The overall ratio between pollinating bees and natural enemies (predatory and parasitic wasps and cleptoparasitic cuckoo bees) was 8.8:2.3.
Table 1. List of bees and wasps collected during field survey at Baghdad-ul-Jadeed campus of The Islamia University of Bahawalpur, Pakistan (September to November, 2019).
Ceratina sp. visited the maximum number (12) of plant species followed by Pseudapis sp. 1 (9), Lasioglossum sp. (8) and Pseudapis sp.2 (7). The minimum interaction with flowering plants was recorded in case of Amegilla sp. (2) and Sphecodes sp. (2). The maximum number (5) of bee species visited Leucaena leucocephala, Trianthema portulacastrum and Carthamus oxyacantha while the minimum number of species (2) visited Calotropis procera (Table 2).
Table 2. List of plant species visited by bee species at Baghdad-ul-Jadeed campus of The Islamia University of Bahawalpur, Pakistan (September to November, 2019).

4. Discussion
In the present study, the bees were more abundant than the wasps. A total of ten bee species in three families and nine wasps in four families were recorded. We surveyed from September to November and found that bees were more abundant than wasps. This finding is supported by a recent study by Sajjad et al. (2019), who reported that bees remained more abundant than wasps in the study area throughout the year. The abundance ratio of bees and wasps vary with climates and regions, i.e. bees are dominant over wasps in sub-tropical regions while wasps dominate over bees in tropical regions (Abrahamczyk et al., 2011). This is because, in sub-tropical regions, the availability of pollen and nectar resources is higher than the tropical regions. On the other hand, in tropical regions, besides feeding on bees, wasps also feed on other arthropods (Potts et al., 2003).
In the present study, cuckoo bees belonged to genus Thyreus (Apidae), Euaspis sp. (Megachilidae) and Sphecodes (Halictidae) while parasitic wasps belonged to genus Podalonia (Sphecidae) and Predatory wasps belonged to genus Polistes (Vespidae) and Vespa (Vespidae) and genus Astata (Crabronidae). Bashir et al. (2019) reported some cuckoo bees and parasitic wasps from southern Punjab; they also reported some genera of family Crabronidae. However, genus Astata and Euaspis are being reported for the first time from the study area.
In the present study, the ground nesting bees were dominant over cavity-nesting bees, i.e. 90.09 % and 9.90 %, respectively. Bahawalpur city mostly lies in Cholistan desert gifted with brown sandy loam soil (Matiullah et al., 2004) and a majority of the soil-nesting bees preferred to construct their nests in sandy loam soil (Cane, 1991). Kim et al. (2006) reported that in sub-tropical regions ground nesting bees were higher in abundance than wood nesting bees. Cleptoparasitic bees have a stabilizing role inside bee communities. The profusion and diversity of cleptoparasitic bees concerning non-parasitic bees indicates the overall status of bee community, and their monitoring can play an essential role in evaluating bee communities (Sheffieled et al., 2013)
Almost 70% of the 30,000 known bee species make a nest in the ground (Cope et al., 2019) Appropriateness of nesting site features for ground-nesting bees is prejudiced by soil moisture, soil texture, slope, depth, vegetation cover, and availability of floral resources (Cane, 2001). Foraging behaviours to collect leaves, resins and mud for segregation of nests also affect the habitat preference (O’Toole and Raw, 1991). However, factors affecting the reproductive success of ground-nesting bees over cavity-nesting bees remain unclear (Morato and Martins, 2006).
In the present study, Ceratina sp. visited the maximum number of plant species followed by Lasioglossum sp. and Pseudapis sp. The minimum interaction with flowering plants was recorded in case of Amegilla sp. and Sphecodes sp. A previous study also suggests similar findings in the study area, i.e. Ceratina and Lasioglossum species visited the maximum number of flowering plants in the study area than Amegilla sp. (Saeed et al., 2012).
In the present study, the maximum number of bee species visited L. leucocephala, T. portulacastrum and C. oxyacantha while the minimum number of bee species visited C. procera. Leucaena leucocephala is a tree with high socioeconomic importance as it is used as fodder for livestock, and the wood is used as fuel (Ngongolo et al., 2014). The brush shaped pale flowers of L. leucocephala are visited by an array of generalist pollinators (Gibson et al., 1989). Ngongolo et al. (2014) reported Apis mellifera, Ceratina and two species of Megachile as essential pollinators of L. leucocephala in Tanzania. Trianthema portulacastrum on the other hand, is a critical weed and act as a vital source of honeybee forage even at the critical time of floral scarcity in India and Pakistan (Dalio, 2015; Sajjad et al. 2019). Carthamus oxyacantha is another noxious weed of agricultural lands and other semi-natural landscapes. Although the flowering ends in early autumn it attracted several bee species in this study.
Flowers of C. procera are melittophilous (Sobrinho et al., 2013) however, few bees visited because nectar is concealed in an almost fully enclosed chamber surrounded by thick tissue with a tiny opening (Wantrop, 1974). Eisikowitch (1986) reported that C. procera was visited only by two species of Xylocopa. Irshad and Stephen (2013) also noticed the minimum number of bee visits on a Calotropis species.
5. Conclusion
The present study gives a brief overview of the assemblage of ground and cavity inhibiting pollinating bees and their natural enemies during the autumn season under sub-tropical climatic conditions. Ground nesting bees were more abundant than cavity-nesting bees. Only a few bee species were much higher in abundance while many species were much lower in abundance. In terms of their abundance, wasps appear to be the more destructive natural enemies of pollinating bees than cleptoparasitic cuckoo bees. The plant species holding the maximum number of bee species interactions can be used to conserve bees in this part of the year, i.e. L. leucocephala, T. portulacastrum and C. oxyacantha. The overall ratio between pollinating bees and natural enemies (8.8:2.3) suggests study area as a balanced ecosystem for the overall process of pollination. Future studies should focus on in-depth tri-trophic interactions of flowering plants, bees and their natural enemies with a viewpoint of quantitative food web predictors and their seasonal turnover as well.
Competing Interest Statement: The authors declare that there are no competing interests.
Author’s Contribution: Aziz-ur-Rehman and Ammad Ahmad undertook the research work while Asif Sajjad and Waseem Akram contributed in the write-up.
Acknowledgements: This research was financed by the Agriculture Linkage Program (ALP) of Pakistan Agriculture Research Council (PARC) under project “Conservation of native bees through ecosystem approach for enhanced crop pollination”.
References
Abrahamczyk, S., J. Kluge, Y. Gareca, S. Reichle and M. Kessler. 2011. The influence of climatic seasonality on the diversity of different tropical pollinator groups. PLoS One 6(11): e27115.
Akram, W., Sajjad, A., Ali, S., Farooqi, M.A., Mujtaba G., Ali, M. and Ahmad, A. 2019. Pollination of Grewia asiatica (Malvaceae) by Megachile cephalotes (Hymenoptera: Megachilidae): Male vs. Female Pollination. Sociobiology. 66(3): 467-474.
Ali, M., Saeed, S. and Sajjad A, Bashir, M.A. 2014. Exploring the best native pollinators for pumpkin production in Punjab Pakistan. Pakistan J. Zool. 46(2): 531-539.
Bashir, M.A., S. Saeed, A. Sajjad, K.A. Khan, H.A. Ghramh, M.A. Shehzad, H. Mubarak, N. Mirza, S. Mahpara, M.I.A. Rehmani and M.J. Ansari. 2019. Insect pollinator diversity in four forested ecosystems of southern Punjab, Pakistan. Saudi J. Biol. Sci. 26(7): 1835-1842.
Cane, J.H. 1991. Soils of Ground-Nesting bees (Hymenoptera: Apoidea): texture, Moisture, Cell depth and Climate. J. Kansas Entomol. Soc. 64(4): 406-413.
Cane, J.H. 2001. Habitat fragmentation and native bees: a premature verdict? Conserv. Ecol. 5(1): 3.
Cane, J.J.H. 2005. Bees’ needs challenged by urbanization. Nature in Fragments – Urban Sprawl. Columbia University Press, New York. p. 109–124.
Cope, G., J.W. Campbell, S.M. Grodsky and J.D. Ellis. 2019. Evaluation of nest-site selection of ground-nesting bees and wasps (Hymenoptera) using emergence traps. Can. Entomol. 151: 260-271.
Dalio, J.S. 2015. Foraging behaviour of Apis mellifera on Trianthema portulacastrum. J. Entomol. Zool. Stud. 3(2): 105-108.
Eisikowitch, D. 1986. Morpho-ecological aspects on the pollination of Calotropis procera (Asclepiadaceae) in Israel. Plant Syst. Evol. 152(3-4): 185-194.
Gibson, G.L., A.R. Griffin and A.C. Matheson. 1989. Breeding tropical trees: population structure and genetic improvement strategies in clonal and seedling forestry. Oxford Forestry Institute, Oxford, UK. p. 218-226.
Goulson, D. 2003. Conserving wild bees for crop pollination. J. Food Agric. Environ. 1: 142-144.
Hausmann, S.L. J.S. Petermann. 2016. Wild bees as pollinators of city trees. Insect Conserv. Diver. 9(2): 97-107.
Holzschuh, A., I. Steffan-dewenter and T. Tscharntke. 2008. Agricultural landscapes with organic crops support higher pollinator diversity. Oikos 117(3): 354–361.
Irshad, M. and E. Stephen. 2013. Value of insect pollinators to agriculture of Pakistan. Int. J. Agron. Agric. Res. 3: 14-21.
Kareiva, P. and U. Wennergren. 1995. Connecting landscape patterns to ecosystem and population processes. Nature 373(6512): 299-302.
Kim, J., N. Williams and C. Kremen. 2006. Effects of cultivation and proximity to natural habitat on ground-nesting native bees in California sunflower fields. J. Kansas Entomol. Soc. 79(4): 309-320.
Kremen, C., N.M. Williams and R.W. Thorp. 2002. Crop pollination from native bees at risk from agricultural intensification. Proc. Natl. Acad. Sci. USA, 99: 16812-16816.
Matiullah, A. Ahad, S. Ur Rehman, S. Ur Rehman and M. Faheem. 2004. Measurement of radioactivity in the soil of Bahawalpur Division, Pakistan. Radiat. Prot. Dosim. 112(3): 443-447.
Michener, C.D. 2000. The bees of the world (Ed. 1st). John Hopkins University Press, Baltimore.
Michener, C.D. 2007. The bees of the world (Ed. 2nd). John Hopkins University Press, Baltimore.
Morato, E.F. and R.P. Martins. 2006. An overview of proximate factors affecting the nesting behavior of solitary wasps and bees (Hymenoptera: Aculeata) in preexisting cavities in wood. Neotrop. Entomol. 35(3): 285-298.
Ngongolo, K., S. Mtoka, A. Mahulu and A. Sigala. 2014. The Leucaena leucocephala floral visitors, pollinators and their predators in the restored Wazo Hill Quarry, Tanzania. Entomol. Appl. Sci. Lett. 1(3): 36-42.
Ollerton, J., R. Winfree and S. Tarrant. 2011. How many flowering plants are pollinated by animals? Oikos 120: 321-326.
O’neill, K.M. 2008. Apoid wasps (Hymenoptera: Apoidea: Spheciformes). In: Capinera J.L. (eds) Encyclopedia of Entomology. Springer, Dordrecht.
O’Toole, C. and A. Raw. 1991. Bees of the World. Blandford Press.
Perfecto, I. and J. Vandermeer. 1996. Microclimatic changes and the indirect loss of ant diversity in a tropical agroecosystem. Oecologia 108: 577–582.
Polis, G.A., W.B. Anderson and R.D. Holt. 1997. Toward an integration of landscape and food web ecology: the dynamics of spatially subsidized food webs. Annu. Rev. Ecol. Syst. 28(1): 289-316.
Potts, S.G., B. Vulliamy, A. Dafni, G. NéEman and P. Willmer. 2003. Linking bees and flowers: How do floral communities structure pollinator communities? Ecology 84: 2628-2642.
Potts, S.G., J.C. Biesmeijer, C. Kremen, P. Neumann, O. Schweiger and W.E. Kunin. 2010. Global pollinator declines: trends, impacts and drivers. Tree 25: 345-353.
Saeed, S., S.A. Malik, K. Dad, A. Sajjad and M. Ali. 2012. In search of the best native pollinators for bitter gourd (Momordica charantia L.) pollination in Multan, Pakistan. Pakistan J. Zool. 44(6): 1633-1641.
Sajjad, A. and S. Saeed. 2010. Floral host plant range of syrphid flies (Syrphidae: Diptera) under natural conditions in southern Punjab, Pakistan. Pakistan J. Bot. 42(2): 1187-1200.
Sajjad, A., M. Ali, S. Saeed, M.A. Bashir, I. Ali, K.A. Khan, H.A. Ghramh and M.J. Ansari. 2019. Yearlong association of insect pollinator, Pseudapis oxybeloides with flowering plants: Planted forest vs. agricultural landscape. Saudi J. Biol. Sci. 26(7): 1799-1803.
Sajjad, A., S. Saeed and S. Burhan-u-din. 2012. Yearlong association of butterfly populations with flowering plants in Multan, Pakistan. Pakistan Entomol. 34(2): 105–110.
Sheffield, C.S., A. Pindar, L. Packer and P. Kevan. 2013. The potential of cleptoparasitic bees as indicator taxa for assessing bee communities. Apidologie. 44(5): 501-510.
Sobrinho, M.S., G.M. Tabatinga, I.C. Machado and A.V. Lopes. 2013. Reproductive phenological pattern of Calotropis procera (Apocynaceae), an invasive species in Brazil: annual in native areas; continuous in invaded areas of caatinga. Acta Bot. Bras. 27(2): 456-
Wantrop, H.E. 1974. Calotropis gigantea (Asclepiadaceae) and Xylocopa tenuiscapa (Hymenoptera, Apidae). studies in flower morphology and pollination biology. Sven. Bot. Tidskr. 68: 25-32.
Zameer, S.U., Fazal, M.I, Bilal, M. and Sajjad, A. (2017). Foraging behavior of pollinators leads to effective pollination in radish Raphanus sativus L. Asian J. Agri. Biol. 5(4): 221-227.