

Journal of Environmental and Agricultural Sciences (JEAS). Nasir and Toth, 2021. Volume 23(3&4): 8-16
Open Access – Research Article
Effect of Drought Stress on Morphology, Yield, and Chlorophyll Concentration of Hungarian Potato Genotypes
Muhammad Waqar Nasir 1,*, Zoltan Toth 1,*
1Institute of Agronomy, Hungarian University of Agriculture and Life Sciences, Georgikon Campus, Keszthely, Hungary
Abstract: Different morphological traits have been studied to determine the drought tolerance of potato genotypes. In this study, a greenhouse experiment was conducted to investigate the effect of drought stress on morphology and yield with a focus on chlorophyll concentration (Chl) of two Hungarian potato genotypes (Hopehely and Demon) varying in drought tolerance. Two potato genotypes (Hopehely and Demon) were grown under two moisture levels i.e. 80% water holding capacity (control) and 50% water holding capacity (stressed). Results revealed that the drought-tolerant genotype (Hopehely) showed significantly higher chlorophyll concentration at each growth stage and produced higher yield under control as well as drought conditions. Drought stress significantly reduced chlorophyll concentration in both genotypes. Tuber yield positively correlated with chlorophyll concentration and specific leaf weight. Chlorophyll concentration varied significantly within the canopy of plants in all treatments. The highest chlorophyll concentration was recorded for the 3rd compound leaf, top leaflet, and top measuring point on the leaflet. Morphological growth of Demon was more sensitive to drought stress compared to Hopehely. This study showed that drought-tolerant genotypes produce higher amount of chlorophyll and maintain their chlorophyll concentration for a longer period even under drought stress.
Keywords: Solanum tuberosum; compound leaves; chlorophyll concentration; SPAD-502; water shortage stress,
*Corresponding author: Muhammad Waqar Nasir: nasir.muhammad.waqar@gmail.com; Zoltan Toth: toth.zoltan@uni-mate.hu
Cite this article as
Nasir, M.W., and Z. Toth. 2021. Effect of drought stress on morphology, yield, and chlorophyll concentration of Hungarian potato genotypes. Journal of Environmental & Agricultural Sciences. 23(3&4): 8-16. [View Full-Text] [Citations]

Copyright © Nasir and Toth 2021. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium provided the original author and source are appropriately cited and credited.
Similar Articles Published in JEAS
- Zaib et al., 2022. Water Stress in Crop Plants, Implications for Sustainable Agriculture: Current and Future Prospects. Journal of Environmental & Agricultural Sciences. 24: xx-xx. [Abstract] [View Full-Text]
- Zahalan, R. and M.M. Alzoubi. 2021. Effect of Organic Physical Soil Amending on Deficit Irrigation Efficiency of potato (Solanum Tuberosum L.). Journal of Environmental & Agricultural Sciences. 23(1&2): 11-18. [ View Full-Text ] [Citations]
- Ahmad, I., S.M.A. Basra, S. Hussain, S.A. Hussain, Hafeez-ur-Rehman, A. Rehman and A. Ali. 2015. Priming with ascorbic acid, salicylic acid and hydrogen peroxide improves seedling growth of spring maize at suboptimal temperature. Journal of Environmental & Agricultural Sciences. 3:14-22. [ ] [View Full-Text] [Citations]
1. Introduction
Water is one of the most critical factors limiting crop productivity (Askari-Khorasgani et al. 2021; Dar et al. 2021). Drought affects plant growth in multiple ways depending upon the duration, intensity, and frequency of drought. Its effect is highly linked with the plant developmental stage (Pinheiro and Chaves, 2011).
The origin of cultivated potato (Solanum tuberosum), is traced back to Latin America. Now it is successfully grown under diverse agroclimatic conditions throughout the world (Bethke et al. 2019; de Haan and Rodriguez, 2016; Saeed et al., 2020). Potato is the 4th most important crop in the world. Due to its rapid increase in consumption, its contribution to regional and global food security is briskly increasing (Jennings et al. 2020). Consequently, sustainable, potato production can be critical to food security and poverty eradication in different regions, and in achieving Sustainable Development Goals (Sing et al., 2020; Wijesinha-Bettoni and Mouillé, 2019).
Potato production is predicted to decrease under changing climate conditions, including drought (Scott et al. 2019; Djaman et al., 2021). Potato developmental stages can be divided into five stages including plant establishment, stolon initiation, tuber initiation, tuber bulking, and maturity stage (Obidiegwu et al. 2015).
Drought can affect potato yield by affecting vegetative growth such as plant height, number, and size of leaves produced by the plant (Deblonde and Ledent, 2001), or by affecting the photosynthesis in leaves by chlorophyll reduction or leaf area index reduction, or leaf area duration reduction. Besides vegetative growth, drought has been reported to affect the reproductive stage of potatoes by shortening the growth cycle (Kumar et al. 2007), or by reducing the size (Schafleitner et al. 2009) and/or numbers of tubers (Eiasu et al. 2007) produced by plants. Moreover, drought can also affect the quality of tubers produced (Ekanayake and Midmore, 1989; Jefferies, 1995).
Potato is a water-efficient crop (Sun et al. 2015) yet considered sensitive to drought stress (Schafleitner et al. 2009), which can be attributed to its shallow root system (Quandahor et al., 2021). Several canopy characteristics such as type of canopy (Aliche et al. 2018), number of leaves (Luitel et al. 2015), and leaf area index (Zarzyńska et al. 2017; Wadas, 2021) have been studied to determine drought tolerance of potato cultivars. Green leaves are the main organ of productivity in terrestrial plants. The presence of chlorophyll in leaves makes them the site of photosynthesis. Chlorophyll content in leaves is used to predict the photosynthetic capacity of the plant, physiological condition, and health of plants (Wright et al. 2004; Zhao et al. 2016). Chlorophyll pigments harness light energy and convert it into chemical energy. The concentration of photosynthetic pigment in the leaves and their concentration in leaves is affected by the nitrogen status of plants, water stress, and other plant stresses (Dai et al. 2009). Chlorophyll concentration may vary among specific parts of the foliage because of the plant growth stage, leaf thickness, leaf position, and the measuring points on the leaf (Ata-Ul-Karim et al. 2016). However, very little research has been reported to understand the effect of chlorophyll concentration variation on potato yield under drought stress.
This study was conducted to observe the morphological variations among Hungarian genotypes under drought stress conditions. This study also aimed to determine the chlorophyll concentration variance in potato plants under drought stress and to determine if there is any relation between chlorophyll concentration and drought tolerance of potatoes.
2. Materials and Methods
2.1. Plant materials and growth conditions
The experiment was conducted in the greenhouse of the Georgikon Campus, Hungarian University of Agriculture and Life Sciences (MATE), Keszthely, Hungary. Two genotypes – Demon and Hopehely – were exposed to two different water levels i.e. 80% water holding capacity (WHC) and 50% WHC. Randomized complete block design in the factorial arrangement was used with 16 replications.
Seed tubers of Demon and Hopehely were obtained from Potato Research Centre, Keszthely, Hungary. Soil and peat mixture (1:1 by weight) was used as growing media in 50 kg soil-bearing pots. Peat of Baltic origin was brought from Latvia and soil was collected from “A” horizon of a Eutric cambisol soil having a sandy clay loam texture from the research farm area of Georgikon Campus MATE. After sieving the soil and peat through a 10 mm sieve; a homogenized mixture was obtained by mixing them in a cement mixer.
Three tubers were sown in each pot. The moisture level was maintained at 80% WHC until the completion of germination (18 DAS) to ensure 100% germination. The gravimetric method was used to determine the water holding capacity of growing media. Pot weight at 80% WHC and 50% WHC was calculated and maintained from germination completion to harvesting. Drought was induced at germination completion (18 DAS). Random pots were weighed weekly to maintain desired water levels in the pots. Thinning was performed at germination completion to maintain 2 plants per pot.
2.2. Observations
2.2.1 Physiological observations
For physiological observations, 4 replications of each genotype at both water levels were harvested at four growth stages: 1. Tuber initiation stage (36 DAS); 2. Flowering stage (54 DAS); 3. Tuber bulking stage (72 DAS); Senescence (90 DAS). At each harvesting data regarding plant height (cm), the number of leaflets per plant, average leaflet weight (g), leaf area index, specific leaf weight (g cm-2), leaf area duration (days), nitrogen percentage in foliage (N%) and tuber yield (g plant-1) was recorded.
Average leaflet weight (ALW) and specific leaflet weight (SLW) were measured by the following formula:

Leaf area index (LAI) and leaf area duration (LAD) were measured at each harvesting by using the following formula:

Where t1 and t2 are the time of first and second sampling and LAI1 and LAI2 are leaf area index at t1 and t2 respectively.
Nitrogen percentage in the foliage was determined by using an elemental analyzer. Leaf and stems were sun-dried followed by oven drying. The dried foliage was mechanically grounded to dust size particles and 100 mg samples were weighed in tin containers (8 × 55 mm) that were then shaped in the form of little round balls. The samples were then delivered to an elemental analyzer using 96 wells plates to determine nitrogen percentage in foliage.
2.2.2. Chlorophyll determination
Chlorophyll content per unit area was determined using Soil Plant Analysis Development (SPAD) 502. The readings were recorded in SPAD units. SPAD values on the top leaflet, 1st side leaflets, and 2nd side leaflets of the 3rd, 4th, and 5th compound leaf from the apex, and at 3 points (top, middle and basal) within a leaflet were taken weekly starting after tuber initiation stage (44 DAS). Three readings were taken for each point and then averaged; hence, 81 readings were taken from each plant (Fig. 1).
2.3. Statistical analysis
SPSS/PASW Statistics for Windows, version 18 (SPSS Inc., Chicago, IL, USA) was used for statistical analysis. Recorded data were subjected to factorial analysis at a 5% significance level followed by Fisher’s Least Significant Difference (LSD) post hoc test to recognize specific differences among treatments. Kendall’s rank correlation was performed to govern the association between variables. A p < 5% was considered significant.
3. Results
At the tuber initiation stage, genotypes showed a significant effect on all foliage characteristics while drought stress didn’t show any significant effect on foliage development of either genotype except plant height of demon. Demon in comparison to Hopehely produced significantly taller plants, a greater number of leaflets, and higher leaf area index (LAI) under control as well as drought conditions; while Hopehely produced higher average leaf weight (ALW), specific leaf weight (SLW), and foliage nitrogen percentage (N%) under control as well as drought condition (Table 1).

Fig. 1. Compound leaf, leaflet on the compound leaf, and measuring point on the leaflet of potato used for chlorophyll measurement (Adopted from Li et al., 2012).
Table 1. Effect of genotypes and drought stress on growth and yield of potatoes at various growth stages

* p<0.05. NOL- number of leaflets per plant; ALW- average leaf weight (g); LAI- leaf area index; SLW- specific leaf weight (g cm-2); LAD- leaf area duration (days); N%- nitrogen percentage in foliage; TY- tuber yield (g).
Similar results were observed at the flowering stage where genotypes varied significantly in their foliage development except LAI for which no significant difference was observed and drought stress significantly reduced the plant height of Demon. At the tuber bulking stage, significant differences were observed between genotypes for plant height, number of leaflets, ALW, N%, and tuber yield. Demon produced significantly taller plants and a greater number of leaflets while Hopehely produced significantly higher ALW, N%, and tuber yield. Besides plant height, drought stress significantly reduced the number of leaflets in Demon at the tuber bulking stage (Table 1).
Table 2. Foliage characteristics of potato genotypes at different growth stages under different water levels

Means followed by different letters in the same column are significantly different (5% probability level); NOL- number of leaflets per plant; ALW- average leaf weight (g); SLW- specific leaf weight (g cm-2); WHC, water holding capacity
Table 3. Chlorophyll concentration variation in the canopy of potato genotypes in response to drought stress
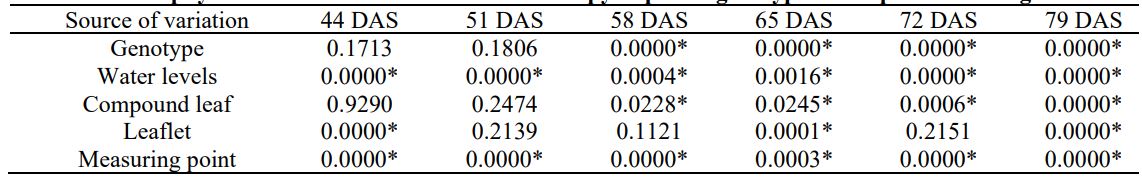
*p<0.05, DAS – Days after sowing
Table 4. Genotypic variation of potato leaf chlorophyll concentration, in response to drought stress

Means followed by different letters in the same column are significantly different at the 5% probability level. WHC- water holding capacity, DAS, days after sowing.
At senescence, significant variation between genotypes was only observed for the number of leaflets and ALW where Demon produced more leaflets and Hopehely produced higher ALW. However, drought stress significantly affected all foliage characteristics of Demon except LAI, while, no significant effect of drought stress was observed on Hopehely (Table 2).
Variance analysis showed that chlorophyll content varied significantly between genotypes after the flowering stage (58 DAS) while drought stress showed a significant effect on chlorophyll content throughout the experiment. A significant difference was observed for chlorophyll content among leaves, leaflets, and measuring points on leaflets (Table 3).
Table 5. Chlorophyll concentration in different foliage parts of Demon at different water levels

Means followed by different letters in the same column are significantly different (5% probability level). W1- 80% water holding capacity (WHC); W2- 50% WHC; L3, L4, L5- 3rd, 4th, 5th compound leaf from the apex respectively; LL1, LL2, LL3; top, first and second leaflet respectively; M1, M2, M3- top middle and bottom measuring point on the leaflet respectively.
Table 6. Chlorophyll concentration in different foliage parts of Hopehely at different water levels
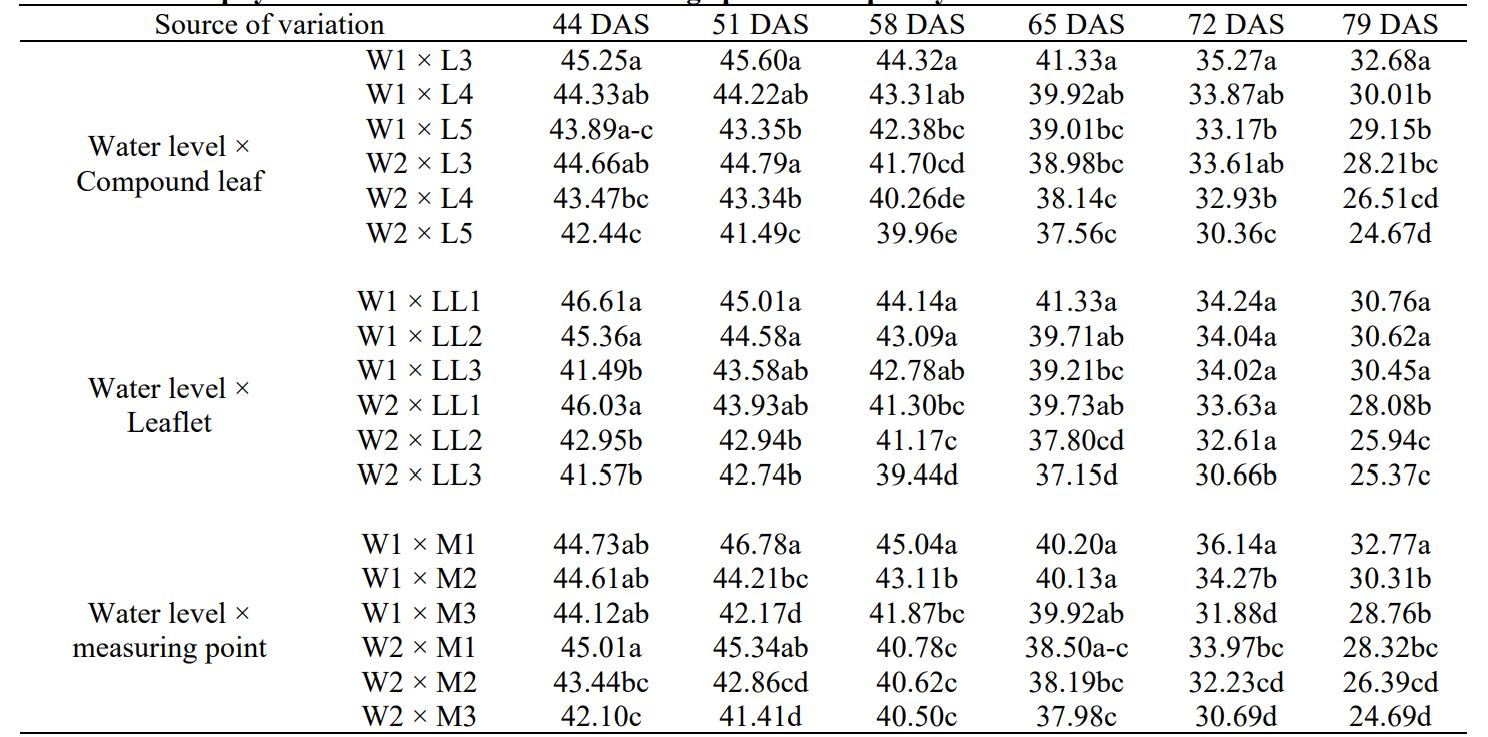
Means followed by different letters in the same column are significantly different (5% probability level). W1- 80% water holding capacity (WHC); W2- 50% WHC; L3, L4, L5- 3rd, 4th, 5th compound leaf from the apex respectively; LL1, LL2, LL3; top, first and second leaflet respectively; M1, M2, M3- top middle and bottom measuring point on the leaflet respectively.
Table 7. Kendall’s Correlations analysis of chlorophyll concentration with tuber yield and foliage characteristics
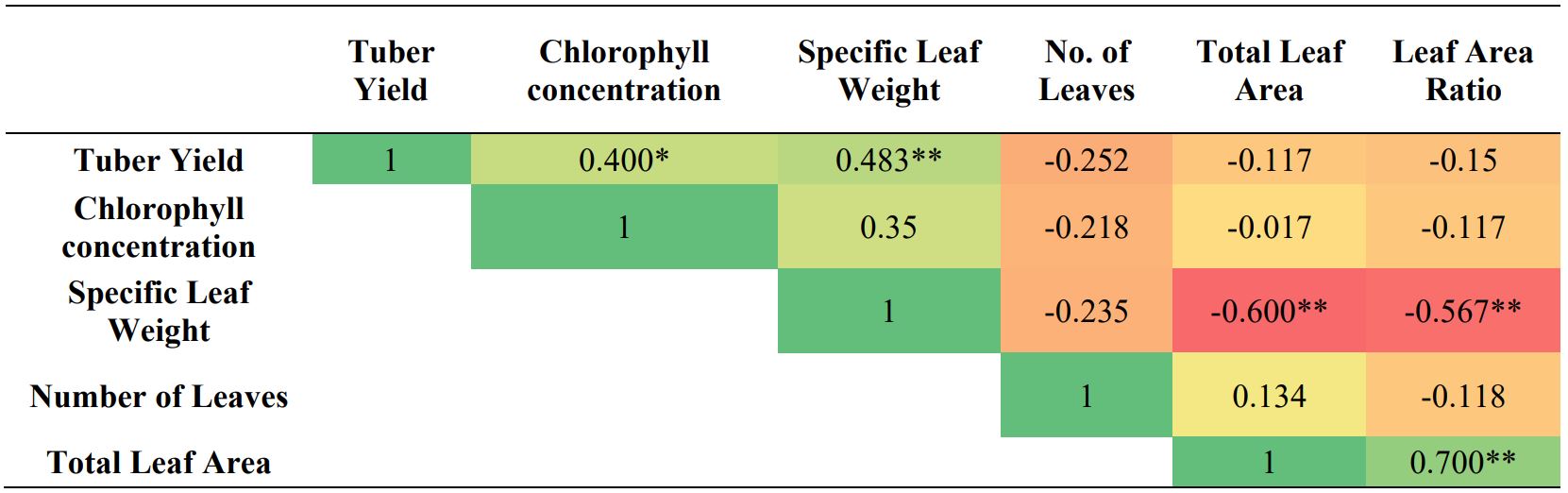
*p<0.05, **p<0.01
In foliage, significantly higher chlorophyll content was observed for 3rd leaf, 1st leaflet, and top measuring point for both genotypes under control as well as drought condition. Both genotypes produced maximum chlorophyll content at tuber bulking that started reducing afterward. In Demon, sharp reduction takes place after flowering, however, Hopehely maintains its chlorophyll content longer and gradual reduction occurs near senescence. In both genotypes, chlorophyll reduction is first observed in lower compound leaves and leaflets (Table 5 and 6). Correlation analysis showed that tuber yield positively correlated to chlorophyll content and specific leaf weight of leaves. It also showed that chlorophyll content was not affected by other foliage characteristics such as specific leaf weight, specific leaf area, or leaf area ratio (Table 7).
4. Discussion
Potato genotype Hopehely produced significantly higher chlorophyll content compared to Demon. It can be due to variation in the foliage development of genotypes. Hopehely produced fewer but heavier leaves with significantly higher average leaf weight (ALW), specific leaflet weight (SLW), nitrogen percentage in foliage (N%), and produced significantly higher tuber yield. Larger and heavier leaves have a great boundary layer thickness that helps in heat exchange, reduces water loss, and maximizes photosynthetic activity by maintaining relative water content (Banik et al. 2016) thus increasing chlorophyll content. On the other hand, Demon produced significantly taller plants with more leaflets, hence, higher LAI throughout the experiment. A large number of leaflets in Demon can be due to the availability of more internodes on taller plants (Tadesse et al. 2001) or due to smaller leaflets size as a negative correlation exists between leaf size and leaf numbers (Westoby and Wright, 2003). A positive correlation was also observed between chlorophyll content and SLW. It shows that plants with smaller and lighter leaves produced less chlorophyll content per unit area as compared to larger and heavier leaves.
Results also highlighted that chlorophyll content varies significantly with in the same plant. In both genotypes, chlorophyll content decreases from top to bottom. It can be due to the lower light intensity received by lower leaves and base points of the same leave due to the shading effect. These results are in line with (Björkman, 1968; Ashworth and Svec, 1974) who reported significantly lower chlorophyll content per unit area in leaf base. Plants reached the highest chlorophyll content before flowering as they completed vegetative growth which shows that a high part of absorbed nitrogen was used in chlorophyll production until flowering. Chlorophyll content started decreasing after flowering as plants start assimilating reserves for tuber production (Argenta et al. 2004; Reynolds et al. 2005). Moving towards senescence lower leaves lost chlorophyll content earlier than upper leaves.
Besides genotype, drought stress also showed a significant effect on foliage development and chlorophyll content of genotypes, particularly Demon. Plant height, the number of leaflets, LAI, LAD, and N% were significantly reduced in Demon, however, no significant difference was observed in foliage development of Hopehely. It shows higher drought tolerance of Hopehely that produced higher tuber yield under stress conditions as compared to Demon. These results are in line with (Kebede et al. 2019) who showed genotypic differences in the ability to maintain leaf expansion with increasing drought stress. However, drought stress significantly reduced per unit chlorophyll content of both genotypes throughout the experiment can be the reason for the significant reduction of tuber yield in Hophely under drought stress. It shows that drought stress causes a reduction in per unit chlorophyll content in drought-sensitive as well as drought-tolerant plants.
5. Conclusion
Potato genotype Hopehely produced higher tuber yield under control as well as drought conditions. Hopehely produced the higher chlorophyll content and specific leaf weight under stress conditions. On the other hand, Demon was highly affected by drought stress. This was shown by the reduction in chlorophyll content, foliage nitrogen percentage, leaf area index, number of leaves, and specific leaf weight. Therefore, Hopehely can be used to obtain high yields particularly in areas of water shortage. Drought stress affects chlorophyll concentration in drought-tolerant as well as sensitive genotypes. Although, tuber yield and chlorophyll content correlate positively, yet, it is important to identify the foliage area that truly represents the chlorophyll concentration of the whole plant. In the present study, we observed significant chlorophyll concentration variation in leaves of both genotypes with the top leaflet of 3rd compound leaf from the apex showing the highest chlorophyll concentration. Further studies to determine the relationship between chlorophyll concentration and tuber yield can help in the selection of suitable drought-tolerant genotypes.
Competing Interest Statement: The authors declare that they have no competing interests.
List of Abbreviations: Chl, chlorophyll concentration; WHC, water holding capacity; DAS, Days after sowing; ALW, Average leaf weight; SLW, Specific leaf weight; LAI, Leaf area index; LAD, Leaf area duration; NOL, Number of leaves; N%, Nitrogen percentage in foliage; TY, Tuber yield.
Author’s Contribution: Conceptualization, M.W.N. and Z.T.; methodology, M.W.N.; software, M.W.N.; validation, M.W.N., and Z.T.; formal analysis, M.W.N.; investigation, M.W.N.; resources, Z.T.; data curation, M.W.N.; writing—original draft preparation, M.W.N.; writing—review and editing, Z.T.; visualization, M.W.N.; supervision, Z.T.; project administration, Z.T.; funding acquisition, Z.T. All authors have read and agreed to the published version of the manuscript.
Acknowledgment: This research was supported by the Hungarian Government and the European Union, with the co-funding of the European Regional Development Fund in the frame of the Széchenyi 2020 Programme GINOP-2.3.2-15-2016-00054 project.
References
Aliche, E.B., M. Oortwijn, T.P. Theeuwen, C.W. Bachem, R.G. Visser and C.G. van der Linden. 2018. Drought response in field grown potatoes and the interactions between canopy growth and yield. Agric. Water Manag. 206:20–30.
Argenta, G., P.R.F. da Silva and L. Sangoi. 2004. Leaf relative chlorophyll content as an indicator parameter to predict nitrogen fertilization in maize. Ciênc. Rural 34:1379–1387.
Ashworth, E.N. and L.V. Svec. 1974. Variations in photosynthesis, chlorophyll content, and ribulose-l, 5-diphosphate carboxylase activity within tobacco leaves. Can. J. Plant Sci. 54:409–412.
Askari-Khorasgani, O., M. I. A. Rehmani, S. H. Wani and A. Kumar. 2021. Osmotic Stress: An Outcome of Drought and Salinity. In Handbook of Plant and Crop Physiology, 445-464. CRC Press.
Ata-Ul-Karim, S. T., Q. Cao, Y. Zhu, L. Tang, M. I. A. Rehmani and W. Cao. 2016. Non-destructive Assessment of Plant Nitrogen Parameters Using Leaf Chlorophyll Measurements in Rice. Front. Plant Sci. 7: 1829.
Banik, P., W. Zeng, H. Tai, B. Bizimungu and K. Tanino. 2016. Effects of drought acclimation on drought stress resistance in potato (Solanum tuberosum L.) genotypes. Environ. Exp. Bot. 126:76–89.
Bethke, P.C., D.A. Halterman and S.H. Jansky. 2019. Potato germplasm enhancement enters the genomics era. Agronomy 9:575.
Björkman, O. 1968. Carboxydismutase activity in shade-adapted and sun-adapted species of higher plants. Physiol. Plant. 21:1–10.
Dai, Y., Z. Shen, Y. Liu, L. Wang, D. Hannaway and H. Lu. 2009. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 65:177–182.
Dar, J. S., M. A. Cheema, M. I. A. Rehmani, S. Khuhro, S. Rajput, A. L. Virk, S. Hussain, M. A. Bashir, S. M. Alghanem, F. M. Al-Zuaibr, M. J. Ansari and K. Hessini. 2021. Potassium fertilization improves growth, yield and seed quality of sunflower (Helianthus annuus L.) under drought stress at different growth stages. PLOS ONE. 16: e0256075.
de Haan, S. and F. Rodriguez. 2016. Potato origin and production. Advances in potato chemistry and technology. Elsevier. pp.1–32.
Deblonde, P.M.K. and J.-F. Ledent. 2001. Effects of moderate drought conditions on green leaf number, stem height, leaf length and tuber yield of potato cultivars. Eur. J. Agron. 14:31–41.
Djaman, K., S. Irmak, K. Koudahe and S. Allen. 2021. Irrigation management in potato (Solanum tuberosum L.) production: A review. Sustainability. 13: 1504.
Eiasu, B.K., P. Soundy and P.S. Hammes. 2007. Response of potato (Solarium tuberosum) tuber yield components to gel-polymer soil amendments and irrigation regimes. N. Z. J. Crop Hortic. Sci. 35:25–31.
Ekanayake, I.J. and D.J. Midmore. 1989. Root-pulling resistance of potatoes in a drought environment. Am. Potato J. 66:519.
Jefferies, R.A. 1995. Physiology of crop response to drought. Potato ecology and modelling of crops under conditions limiting growth. Springer. pp.61– 74.
Jennings, S.A., A.-K. Koehler, K.J. Nicklin, C. Deva, S.M. Sait and A.J. Challinor. 2020. Global potato yields increase under climate change with adaptation and CO2 fertilisation. Front. Sustain. Food Syst. 4.
Kebede, Z., F. Mekbib, T. Abebe and A. Asfaw. 2019. Evaluation of leaf-water relation traits, as selection criterion for developing drought resistant potato (Solanum Tuberosum L.) Genotypes. Evaluation 6:1–10.
Kumar, S., R.A.M. Asrey and G. Mandal. 2007. Effect of differential irrigation regimes on potato (Solanum tuberosum) yield and post-harvest attributes.
Li, L., Y. Qin, Y. Liu, Y. Hu and M. Fan. 2012. Leaf positions of potato suitable for determination of nitrogen content with a SPAD Meter. Plant Prod. Sci. 15:317–322.
Luitel, B.P., B.B. Khatri, D. Choudhary, B.P. Paudel, S. Jung-Sook, O.-S. Hur, H.J. Baek, K.H. Cheol and R.K. Yul. 2015. Growth and yield characters of potato genotypes grown in drought and irrigated conditions of Nepal. Int. J. Appl. Sci. Biotechnol. 3:513–519.
Obidiegwu, J.E., G.J. Bryan, H.G. Jones and A. Prashar. 2015. Coping with drought: stress and adaptive responses in potato and perspectives for improvement. Front. Plant Sci. 6.
Pinheiro, C. and M.M. Chaves. 2011. Photosynthesis and drought: can we make metabolic connections from available data? J. Exp. Bot. 62:869–882.
Quandahor, P., Y. Gou, C. Lin, J. A. Coulter and C. Liu. 2021. Comparison of root tolerance to drought and aphid (Myzus persicae Sulzer) resistance among different potato (Solanum tuberosum L.) cultivars. Sci. Rep. 11: 628.
Reynolds, M.P., A. Mujeeb-Kazi and M. Sawkins. 2005. Prospects for utilising plant-adaptive mechanisms to improve wheat and other crops in drought-and salinity-prone environments. Ann. Appl. Biol. 146:239–259.
Saeed, B., A. Nawab and S. Rani. 2020. Genotypic variation in micronutrient composition of potato (Solanum tuberosum). J. Environ. Agric. Sci. 22: 64-70.
Schafleitner, R., R. Gutierrez, S. Legay, D. Evers and M. Bonierbale. 2009. Drought stress tolerance traits of potato. Proceedings of the 15th International Symposium of the International Society for Tropical Root Crops (ISTRC), Lima.
Scott, G.J., A. Petsakos and H. Juarez. 2019. Climate change, food security, and future scenarios for potato production in India to 2030. Food Secur. 11:43–56.
Singh, B., P. Raigond, S. Dutt and M. Kumar. 2020. Potatoes for Food and Nutritional Security. Potato. Springer. pp.1–12.
Sun, Y., X. Cui and F. Liu. 2015. Effect of irrigation regimes and phosphorus rates on water and phosphorus use efficiencies in potato. Sci. Hortic. 190:64–69.
Tadesse, M., W.J.M. Lommen, P.E.L. Van der Putten and P.C. Struik. 2001. Development of leaf area and leaf number of micropropagated potato plants. NJAS-Wageningen. J. Life Sci. 49:15–32.
Wadas, W. 2021. Potato (Solanum tuberosum L.) growth in response to foliar silicon application. Agronomy. 11: 2423.
Westoby, M. and I.J. Wright. 2003. The leaf size–twig size spectrum and its relationship to other important spectra of variation among species. Oecologia 135:621–628.
Wijesinha-Bettoni, R. and B. Mouillé. 2019. The contribution of potatoes to global food security, nutrition and healthy diets. Am. J. Potato Res. 96:139–14
Wright, I.J., P.B. Reich, M. Westoby, D.D. Ackerly, Z. Baruch, F. Bongers, J. Cavender-Bares, T. Chapin, J.H. Cornelissen and M. Diemer. 2004. The worldwide leaf economics spectrum. Nature 428:821–827.
Zarzyńska, K., D. Boguszewska-Mańkowska and A. Nosalewicz. 2017. Differences in size and architecture of the potato cultivars root system and their tolerance to drought stress. Plant Soil Environ. 63:159–164.
Zhao, B., Z. Liu, S.T. Ata-Ul-Karim, J. Xiao, Z. Liu, A. Qi, D. Ning, J. Nan and A. Duan. 2016. Rapid and nondestructive estimation of the nitrogen nutrition index in winter barley using chlorophyll measurements. Field Crops Res. 185:59–68.
Zulfiqar, U., S. Hussain, M. Maqsood, M. Ishfaq and N. Ali. 2021. Zinc nutrition to enhance rice productivity, zinc use efficiency, and grain biofortification under different production systems. Crop Sci. 61(1): 739-749.
Join Journal of Environmental and Agricultural Sciences (JEAS)
Interested to join the JEAS Team
Join JEAS as a member Editorial Board see Editors’ Responsibilities
Join JEAS as a member Review Panel Reviewers’ Responsibilities
(send your CV through email at editor.jeas@outlook.com)
JEAS Indexing Journal of Environmental EAS is indexed by reputed indexing services.
Suggest Indexing service/s through email (editor.jeas@outlook.com)
Call for Articles
Submit Your research for publication in the “Journal of Environmental and Agricultural Sciences (JEAS)” through email: editor.jeas@outlook.com
- How to prepare your manuscript before submission
- How to submit your manuscript
- Publication Ethics
- Publication Fee Currently JEAS is publishing manuscripts without publication or processing fee
JEAS Recently Published and Highly Cited Articles
Citation record of JEAS: JEAS Google Scholar page
Follow JEAS Facebook
I went over this website and I conceive you have a lot of great information, saved to fav (:.