

Journal of Environmental and Agricultural Sciences (JEAS). Otiende and Maimba, 2020. Volume 22(1): 1-9
Open Access – Research Article
Endogenous Carbohydrate Content of the Cutting Positions at Time of Severance and IBA Concentration Influence Rooting of Rosa hybrida Rootstocks
Millicent Adhiambo Otiende 1,* , Francis Munyiri Maimba 2
1 University of Kabianga, Kericho, Kenya
2 James Finlay (Kenya) Limited, Kenya
Abstract: Insufficient rooting of cuttings may result in economic losses and limit the use of some rootstocks. Present study was aimed at evaluating the effect of cutting position (top, middle and bottom) and Indole-3-butyric acid (IBA) concentrations (0%, 0.2%, 0.4% and 0.6%) on rooting of Rosa Progress and Natal Briar cuttings. Carbohydrate content of the cutting positions at the time of severance was determined. The experiment was factorial in a completely randomized design (CRD). The root and shoot growth parameters as well as carbohydrate levels in the leaves increased acropetally in both rootstocks. Indole-3-butyric acid (IBA) treatment enhanced root number, total root length and root fresh weight of both rootstocks. Among the IBA treated cuttings 0.4% had higher rooting and survival percentage than 0.2% in Natal Briar. Rosa Progress exhibited higher root number, shooting and survival percentage than Natal Briar though carbohydrate content in the original leaf at the time of severance was low. Carbohydrate content in the leaf of both rootstocks was higher than in the stem and this presumably ensured continuous supply of carbohydrates to the stem base during the rooting process. Auxin (IBA) treatment and high endogenous carbohydrate in the original leaf at the time of cutting severance enhanced rooting especially in Natal Briar.
Keywords: Carbohydrate, Rosa hybrida, IBA, cutting position, rooting.
*Corresponding author: Millicent Adhiambo Otiende: millyotiende@yahoo.com
Cite this article as: Otiende, M.A. and F.M. Maimba. 2020. Endogenous carbohydrate content of the cutting positions at time of severance and IBA concentration influence rooting of Rosa hybrida rootstocks. Journal of Environmental & Agricultural Sciences. 22(1):1-9.
1. Introduction
Rosa hybrida, a sign of love, grace, inspiration and source of enjoyment. Globally it is one of the most famous and popular cut flower, of significant commercial importance. Rootstocks have been used to improve performance of the scion such as vigor, flower yield and quality (Otiende et al., 2015; Amiri et al., 2014), nutrient status (Albacete et al., 2015; Esmaeili et al., 2015), disease resistance (Suchoff et al., 2015; Cantero-Navarro et al., 2016) and response to environmental conditions (Li et al., 2016b; Bhatt et al., 2015). Multiplication of the rootstocks can be done either by seed or vegetative means.
Among the vegetative means, stem cutting is mostly used to maintain clones and genetic purity. The physiological and biochemical characteristics of the stock plant as well as treatment of the cuttings derived therein are important in adventitious root formation.
Rooting potential of the cuttings from a physiologically mature stem may vary depending on their positions on the mother stem (Rakibuzzaman et al., 2018; Otiende et al., 2017; Saifuddin et al., 2013) and the variation has been attributed to changes in endogenous carbohydrate content, mineral nutrients, juvenility factors (Otiende et al., 2017; da Costa et al., 2013; Osterc et al., 2009), endogenous auxin, and other biochemical components, including phenolics, contribute as auxin modulators or rooting co-factors (da Costa et al., 2013; Osterc et al., 2009).
Response of cutting position to rooting may also vary depending on the type of vegetative propagation used. For instance in the self-rooting experiment, apical cuttings yielded significantly higher rooting potential as compared to the cuttings from bottom position of rose rootstock (Otiende et al., 2017). However, in the top grafting experiment, grafts on bottom position cuttings of the same rootstock had better rooting potential than grafts on apical cuttings (Otiende et al., 2016).
In literature adventitious root formation is well known to be directly linked to functions of auxin, produced naturally in the plant or exogenously applied. In any of these cases auxin is important for formation of adventitious roots (Ahkami et al., 2013; Wendling et al., 2015; Gudeva et al., 2017). Moreover, auxin concentration regulates rooting capacity of cuttings (Ahkami et al., 2013). Indole-3- butyric-acid (IBA), biologically stable compound, is the main form of auxin exogenously applied to the cutting to promote rooting and subsequent root system for better vigor of plants. Optimal concentration of IBA is key to stimulate cell division in the vascular cambium (Husen, 2012). Subsequent formation of root primordial or stimulating redistribution and mobilization of some auxin cofactors towards base of the cuttings promotes root formation.
Auxins have been used to stimulate adventitious root formation in a number of horticultural plants and forest trees. The influence of cutting position and application of rooting hormones has been reported in own rooted cuttings of Stevia (Rakibuzzaman et al., 2018), honey bush (Cyclopia spp.) (Mbangcolo et al., 2013), grafted rose cuttings (Otiende et al., 2016) and woody plant species.
Involvement of carbohydrates to adventitious roots formation is mainly as carbon and energy source, essential for cell divisions, formation of root meristems (da Costa et al., 2013). These carbohydrates may already be present in the cutting or accumulated through photosynthesis during the rooting period.
Ample reserve of initial carbohydrates in the cutting is required to ensure sufficient energy supply throughout the rooting period (Ruedell et al., 2013). In an effort to enhance multiplication of rose rootstocks, this study was designed to investigate the effects of IBA concentrations and cutting position on rooting potential of own rooted rose stem cuttings. We hypothesized that the concentration of exogenous auxins differentially affects rooting of different cutting positions of rose rootstocks.
2. Materials and Methods
2.1. Experimental Layout and Treatments
Stem cuttings of two Rosa cultivars, Progress and Natal Briar were obtained from mother stock plants (3 month-old) grown at Finlays Flowers Company, Kericho, Kenya. Uniform vertical shoots (150 cm each) were further divided into three equal positions of top (TPC), middle (MPC) and bottom (BPC) (Otiende et al., 2017). Each stem cutting from each position measured 5-6cm long. These cuttings of two rootstocks (Natal Briar and Rosa Progress) were categorized into 3 cutting positions (TPC, MPC, BPC) and exposed to 4 IBA concentrations (0, 0.2%, 0.4% and 0.6%). These treatments were laid out in a completely randomized design (CRD) with factorial combination and performed in triplicate. Each experimental unit had 20 potted plants.
During planting the basal ends of the cuttings were dipped in appropriate IBA concentrations for 5 seconds before being inserted into jiffy bags containing clean sterile coccus with a PH of 6.5-7.5 and electrical conductivity (EC) of 0.18-0.24mS/cm.
The experiment was set in a greenhouse equipped with a misting and heating system at Harvest Flower Company in Athi River, Kenya. Relative humidity of ≥90%, temperature of 30-35oC (day time) and 22- 24oC (night time) and misting cycles of 10-30 minutes (day time) and 1-2 h (night time) were maintained in the first 2 weeks after planting and then gradually reduced to harden the plants. Light intensity was maintained at 300 watts/m2 throughout the growing period. After 30-days experiment various shoot, leaf and root parameters were recorded using standard procedures, including leaf number, shoot and weights, total root number, length, and weights, shooting, rooting and survival percentages.
2.2. Carbohydrate Analysis
The stem cuttings of randomly selected six unrooted cuttings were collected on the day of planting and stored at -20oC to inhibit enzymatic activity. Each stem cutting was separated into two sections; original leaf and stem bases (1 cm). Collected samples were ground, separately for each treatment, to fine particles and homogenize using Wiley mill (Moullnex AR II, China). Modified high performance liquid chromatography (HPLC) method was adopted to determine soluble carbohydrates (glucose, fructose and sucrose) through chromatographic separation and using collected samples (5g each) (Otiende et al., 2017; RybakChmielewska, 2007).
Ethanol (40ml, 96%) was used to extract carbohydrates from cuttings and subsequent centrifugation (13,800 rpm for 20 minutes). The extracts were then filtered through Whatman No. 2 filter paper then reconstituted with 50% v/v HPLCgrade acetonitrile in water. The analysis by HPLC (CBM-20A, Shimadzu, Japan) was performed under isocratic conditions and room temperature (25±1 °C). Stem sugars, at each cutting position, were identified by using Autosampler (SIL-20A, Shimadzu, Japan) and refractive index detector, by comparing retention times of individual carbohydrates.
2.3. Statistical Analysis
A three-way analysis of variance (ANOVA) was done using GENSTAT statistical package. Mean separation was done using Fisher’s least significant difference (LSD) at 5% significant level.
3. Results
3.1. Effects of cutting position on growth parameters of Rosa hybrida rootstocks
The interaction of rootstock and cutting position was significant and all the shoot and root growth parameters measured increased acropetally in both rootstocks with TPC having higher growth performance than BPC (Table 1).
Table 1. Interactive effect of rootstock and cutting position on shoot and root growth parameters of Rosa hybrida rootstocks
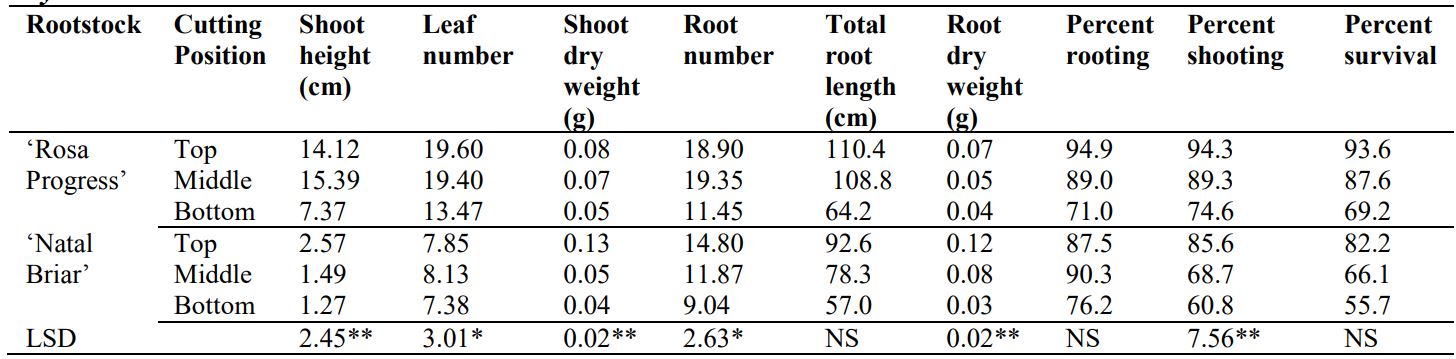
LSD, least significant difference of means; *, significant at p ≤ 0.05; **, significant at p ≤ 0.001; NS, Not significant
3.2. Effects of auxin treatment on growth parameters of Rosa hybrida rootstocks
Auxin treatment significantly promoted the root formation (Table 2) than the control in both rootstocks. Among the IBA treatments, 0.6% IBA exhibited higher root number than 0.2%IBA and 0.4%IBA in Natal Briar. However, IBA application on Rosa Progress could not produce significant effect.
Table 2: Effects of interaction of rootstocks and IBA concentrations on rooting of Rosa hybrida
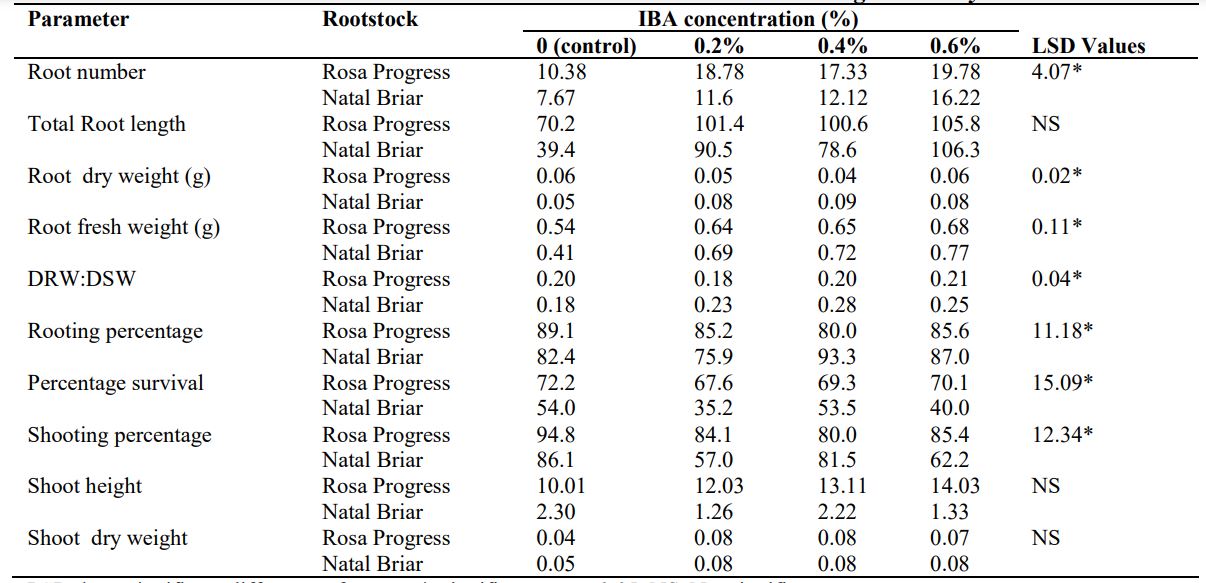
LSD, least significant difference of means; *, significant at p ≤ 0.05; NS, Not significant.
Table 3: Effects of IBA concentration on rooting Rosa hybrida rootstocks

LSD, least significant difference of means; *, significant at p ≤ 0.05; NS, Not significant.
No significant interaction effect of rootstock and auxin was noted for total root length (Table 2), however, the main factor of auxin was significant (Table 3). The total root length increased from the control to 0.2%IBA and 0.6%IBA. The total root length of 0.4% IBA treated cuttings was not significantly different from 0.2%IBA treated cuttings. The control had significantly lower root dry weight (Table 2) than IBA treated cuttings in Natal Briar, however, no significant difference was noted among the IBA treated cuttings. In Rosa Progress, 0.4%IBA exhibited lower root dry weight than 0.6%IBA treated cuttings. Among the IBA treated cuttings, 0.4%IBA had significantly higher rooting percentage than 0.2%treated cuttings but not significantly different from the control and 0.6%IBA treated cuttings in Natal Briar (Table 2).
The root dry weight to shoot dry weight ratio (RDW: SDW) was significantly higher in the auxin treated cuttings than the control. Among the auxin treated cuttings, application of 0.4% auxin concentration had higher RDW: SDW than 0.2%IBA treated cuttings (Table 3). Percent rooting and survival were not significantly different among the treatments in Rosa Progress though, the control had high rooting and survival percentages (Table 2).Percent shooting was significantly higher in the control than 0.4% IBA treated cuttings but not significantly different from the other IBA treated cuttings in Rosa Progress. In Natal Briar, 0.2%IBA treated cuttings exhibited significantly lower percentage shooting and survival than the control but not significantly different from 0.6%IBA treated cuttings (Table 2). The shoot height and shoot dry weight were not affected by the auxin concentration in both rootstocks (Table 2).
Table 4. Interactive effect of rootstock and cutting position on carbohydrate level (%) of the leaf on stem cutting at the time of planting

LSD, least significant difference of means; *, significant at p ≤ 0.05; **, significant at p ≤ 0.001; NS, Not significant
Table 5. Effects of rootstock cultivars and cutting position on carbohydrate level (%) of the stem base of stem cutting at the time of planting
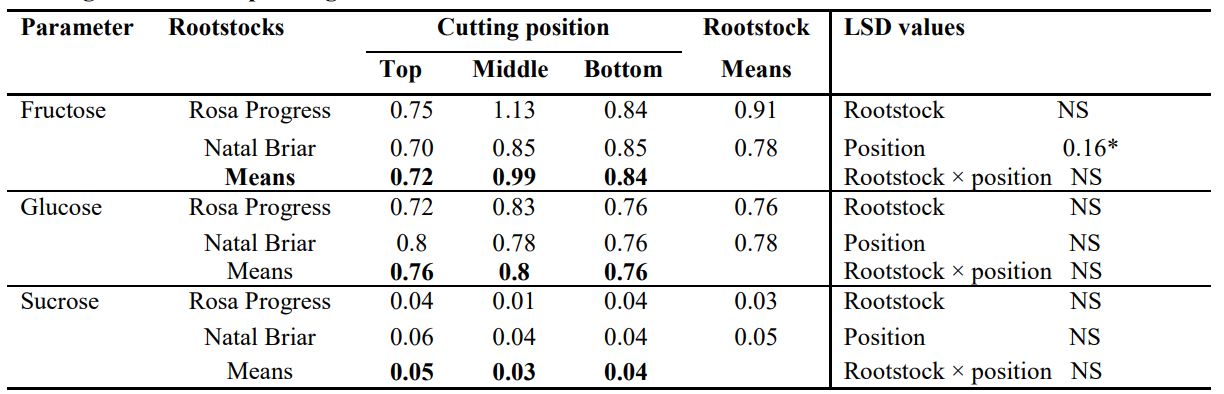
LSD, least significant difference of means; *, significant at p ≤ 0.05; NS, Not significant.
3.3. Carbohydrate levels in the leaf and stem base of the stem cuttings at the time of planting as influenced by rootstock cultivars and cutting position
Rootstock cultivars, cutting position and their interaction produced significant effect on fructose, glucose and sucrose level in the original leaf at the time of planting (Table 4). In case of Natal Briar’, concentrations of fructose and glucose showed significant decreasing trend vertically from TPC to the BPC. Interestingly, sucrose level was significantly higher in MPC of Natal as compared to both TPC and BPC (Table 4). Glucose level in BPC of Rosa Progress was significantly lower than the MPC. However, the glucose level of the TPC was statistically at par with that from the MPC and BPC (Table 4).
The sucrose level significantly decreased from the TPC to the BPC in Rosa Progress. Fructose levels of the three cutting positions were however, not significantly different from each other in Rosa Progress. Irrespective of cutting position, leaves of Natal Briar had significantly higher fructose, glucose and sucrose level than those of Rosa Progress (Table 4). In the stem base of the cuttings, the cutting position significantly influenced the fructose content and the MPC exhibited significantly higher fructose content than the TPC (Table 5). However, the fructose content of the BPC was not significantly different from the other two positions (Table 5). The cutting position however, had no significant effect on the glucose and sucrose contents in the stem base of the cuttings (Table 5).
4. Discussion
4.1. Effects of cutting position on growth parameters of Rosa hybrida rootstocks
The acropetal increase in growth has been associated with juvenility factors (Wendling et al., 2015; Ahkami et al., 2013), endogenous root promoting substances from the terminal buds (Osterc et al., 2016) and high soluble carbohydrates, nitrogen and zinc in the stem cutting (Otiende et al., 2017).
4.2. Effects of auxin treatment on growth parameters of Rosa hybrida rootstocks
Auxin treatment had a promotive effect on rooting potential (root number, fresh and dry weights of roots, RDW: SDW and rooting percentage) of both rootstocks. The promotive effect was more pronounced in Natal Briar than Rosa Progress (Table 2). Auxin is believed to be an effective inducer of adventitious root formation (Pacurar et al., 2014; Rasmussen et al., 2015). Exogenous auxin application stimulates cell differentiation, promotes starch hydrolysis and involve in relocation of nutrients and sugars to the cutting base. All these steps ultimately promote rooting (Atangana et al., 2011). Accumulated auxin concentration, exogenous and endogenous, regulates the divisions of very first root initials (Kochhar et al., 2005).
Application of auxin promotes the rooting of cuttings in roses and Malus spp. (Hartman et al., 2011) and increases the number of developing adventitious roots on cuttings (Ezekiel, 2010). The fine roots produced in stem cuttings are capable of absorbing relatively more water and minerals due to the increment of root surface and root –soil interaction (Stokes et al., 2009). The increase in root length observed in the IBA treated cuttings (Table 3) could be attributed to metabolite accumulation at the site of application of auxins, cell enlargement, enhanced hydrolysis of carbohydrates which provides energy for meristematic tissues of roots (Ezekiel, 2010), synthesis of new proteins and cell division induced by the auxins (Shan et al.,2012). Azad et al., (2016) and Babu et al., (2018) demonstrated that IBA has an important role in the development of adventitious roots, improving quality of roots and increasing root biomass.
The rooting percentage modified significantly depending on concentration of applied hormone (Elhaak et al., 2015; Sevic and Guney, 2013). Among the IBA treated cuttings, 0.4% IBA exhibited significantly higher rooting percentage than 0.2%IBA treated cuttings in Natal Briar (Table 2). Percent rooting and survival were not significantly different among the treatments in Rosa Progress though, the control had high rooting and survival percentages (Table 2). Sevic and Guney (2013) and Asl et al. (2012) also reported comparable results for both the auxin treated cuttings and the control in Melissa officinalis and Bougainvillea species respectively.
Percent shooting was significantly higher in the control than 0.4% IBA treated cuttings but not significantly different from the other IBA treated cuttings in Rosa Progress (Table 2). It is possible that the cuttings of the rootstock Rosa Progress had sufficient amount of endogenous auxins to facilitate adventitious root formation or the concentration of exogenously applied IBA was slightly higher thus slowing adventitious root formation. In Natal Briar, 0.2% IBA treated cuttings exhibited significantly lower percentage shooting and survival than the control but not significantly different from 0.6% IBA treated cuttings. Percentage shooting was dependent on the number of cuttings which produced shoots. Whereas, the survival was based on the number of cuttings that had both roots and shoot.
Root and shoot growth are interrelated as they depend on the photoassimilates from the leaves and hormones especially cytokinins and auxins from the roots and shoot respectively. Both cytokinin synthesized in the roots (Nordstrom et al., 2004) or locally in the node bearing the bud (Xu et al., 2015) regulates the bud outgrowth. After stem decapitation, indole-3-acetic acid (IAA) suppresses the genes involved in synthesis of cytokinins in the node terminates, causing the increased levels of cytokinins (Muller et al., 2015; Xu et al., 2015). Hence the presence of endogenous cytokinin in the stem tissues (Wroblewska, 2013) with the addition of auxin into the basal ends might have eventually promoted the formation of shoot from the cuttings.
Accelerated root formation in Schefflera cuttings, resulted in early onset of axillary bud growth (Hansen and Kristensen, 2006). The promotive effect of auxin on the vegetative growth can be attributed to the enhanced rooting percentage and root growth on the auxin treated cuttings which has higher potential of water and nutrients uptake from the growing medium.
4.3. Carbohydrate levels in the leaf and stem base of the stem cuttings at the time of planting as influenced by rootstock cultivars and cutting position
Being photosynthetically active, the original leaf act as the key source of carbohydrate supply. In addition, it also provides indole-acetic acid, organic nitrogen, vitamins and rooting co-factors, needed for rooting. The carbohydrate (glucose, fructose and sucrose) level increased acropetally in the rootstock Natal Briar with BPC leaves (Table 4) recording lower levels than TPC leaves and this was related to high root-shoot growth responses in TPC (Tables 1). Carbohydrates, being rich source of energy and carbon, promote formation of adventitious roots, mainly by triggering cell division, establishment of the new root meristems and root formation itself (da Costa et al., 2013). The original leaf of single-node soft wood cuttings has a strong effect on survival and rooting success (Thomas and Schiefelbin, 2004) and the growth of the primary shoot and axillary bud of cuttings depends on the assimilate supply from the subtending leaf (Thomas and Schiefelbin, 2004).
In rose plant, ample supply of carbohydrates by the leaf ensures axillary bud outgrowth (Van Labeke et al., 2001). Sucrose promotes auxin export out of the bud which has been associated with bud outgrowth (Barbier et al., 2015; Mason et al., 2014) possibly due to increased levels of cytokinins. This explains the enhanced shoot growth in the TPC.
The stem can be a potential energy source energy or a storage buffer, through the reserves accumulated prior severance, contributing to growth and maintenance of the roots or the primary shoot (Costa and Chall, 2002). Moreover, it also act as sink for photoassimilates from the leaves, mainly due to its respiratory activity (maintenance and growth), storage capacity and little photosynthetic activity. The carbohydrate levels in the cutting tissues is affected by the initial carbohydrate reserves (Druege et al., 2004) and the current carbon assimilation during rooting by photosynthetic activities (Rapaka et al., 2005). At the time of severance, the carbohydrate content of the stem base of the three positions was not significantly different for sucrose and glucose except for fructose that was significantly higher in the MPC than TPC of both rootstock cultivars (Table 5).
The high carbohydrate reserves at the time of severance observed in the leaves located at TPC and MPC of both rootstock cultivars (Table 4). Potentially it will ensure continuous supply of carbohydrates to the stem base during formation of adventitious roots (Otiende et al., 2017), which recorded lower levels of sucrose (Table 5). Leaf sugar concentration at planting time, particularly sucrose, is vital for subsequent formation of adventitious roots in pelargonium cuttings. Because it will ensure carbohydrate supply to the stem bases, site of rooting (Druege et al., 2004; Rapaka et al., 2005). Sucrose is usually hydrolyzed into its constituent monosaccharides (fructose and glucose) by the increased activities of cell wall invertase or decreased activities of vacuolar and cytosolic invertases before it can be utilized in metabolic processes (Kerner et al., 2000; Ahkami et al., 2009).
The acropetal increase in rooting percentage recorded in both rootstock cultivars (Table 1) could be attributed to the high carbohydrate content of the apical leaves at the time of severance in addition to other rooting factors such as mineral nutrition (Otiende et al., 2017) auxins and co factors. The low carbohydrate content of the BPC leaf (Table 4) could be due to declined rate of photosynthesis with increasing leaf age.
5. Conclusion
The carbohydrate content of leaf on the cuttings at the time of severance is important for rooting stem cuttings of the two rootstocks and the bottom position cuttings with low carbohydrate content had the least rooting percentage. Auxin treatment also promoted rooting of the stem cuttings. Further studies can be done to examine the endogenous levels of auxins and rooting co factors that favored rooting in non-IBA treated stem cuttings of Rosa Progress.
List of Abbreviations: ANOVA, Analysis of Variance; BPC, bottom position cutting; DAAD, Deutscher Akademischer Austausch Dienst (German Academic Exchange Service); EC, electrical conductivity; GENSTAT General Statistics software package; HPLC, High Performance Liquid Chromatography; IBA, Indolebutyric acid; NACOSTI- National Commission for Science, Technology and Innovation, ; MPC, middle position cutting; RDW: SDW-Root Dry Weight to Shoot Dry Weight ratio,. TPC, top position cutting.
Competing Interest Statement: The authors declare that there are no competing interests.
Author’s Contribution: M.A.O. and FMM designed, conducted experiment, analyzed data and wrote paper. Acknowledgments: Authors are greatly indebted to Finlay Flowers Limited and Harvest Flowers Company for providing the propagation facilities.
References
Ahkami, A.H., M. Melzer, M. R. Ghaffari, S. Pollmann, M. G. Javid, .F.M. R Shahinnia, M. J. Hajirezaei and U. Druege. 2013. Distribution of indole-3-acetic acid in Petunia hybrida shoot tip cuttings and relationship between auxin transport, carbohydrate metabolism and adventitious root formation. Planta. 238: 499–517.
Ahkami, A.H., S. Lischewski, K.T. Haensch, S. Porfirova, J. Hofmann, H. Rolletschek, M. Melzer, P. Franken, B. Hause, U. Druege and M.R. Hajirezaei. 2009. Molecular physiology of adventitious root formation in Petunia hybrida cuttings: Involvement to wound response and primary metabolism. New Phytol. 181(3): 613– 625.
Albacete, A., C. Andújar, F. Pérez-Alfocea, J. Lozano and M. Asins. 2015. Rootstock-mediated variation in tomato vegetative growth under low potassium or phosphorous supplies. Acta Hortic. 1086: 147– 152.
Amiri, M.E., E. Fallahi and M. Safi-Songhorabad. 2014. Influence of rootstock on mineral uptake and scion growth of ‘golden delicious’ and ‘Royal Gala ’apples. J. Plant. Nutr. 37(1): 16–29.
Asl, M.B., S. Shakueefar and V. Valipour. 2012. Effects of indole-3-butyric acid on the rooting ability of semi hardwood Bouganvillea sp. cuttings. Mod. Appl. Sci. 6(5): 121-123.
Atangana, A.R., M.L.N. Mpeck-Nyemeck, S.X. Chang and D.P. Khasa. 2011. Auxin regulation and function: Insights from studies on rooted leafy stem cuttings of tropical tree species. In: Keller, A.H., Fallon, M.D. (Eds.), Auxins: Structure, Biosynthesis Functions. New York: Nova Science Publishers, Inc. p. 53–66.
Azad, M.S., M.J. Alam, A.S. Mollick and M.A. Martin. 2016. Responses of IBA on rooting, biomass production and survival of branch cuttings of Santalum album L., a wild threatened tropical medicinal tree species. J. Sci. Technol. Environ. Inform. 3(2): 195-206.
Babu, B.H., A. Larkin and H. Kumar. 2018. Effect of plant growth regulators on rooting behavior of stem cuttings of Terminalia arjuna (Roxb). Plant Arch. 18(2): 2159-2164.
Barbier, F., T. Péron, M. Lecerf, M.D. Perez-Garcia, Q. Barrière, J. Rolcík, S. Boutet-Mercey, S. Citerne, R. Lemoine, B. Porcheron, H. Roman, N. Leduc, J. L. Gourrierec, J. Bertheloot and S. Sakr. 2015. Sucrose is an early modulator of the key hormonal mechanisms controlling bud outgrowth in Rosa hybrida. J. Exp. Bot. 66: 2569–2582
Bhatt, R.M., K.K. Upreti, M.H. Divya, S. Bhat, C.B. Pavithra and A.T. Sadashiva. 2015. Interspecific grafting to enhance physiological resilience to flooding stress in tomato (Solanum lycopersicum L.). Sci. Hortic.182: 8–17.
Cantero-Navarro, E., R. Romero-Aranda, R. Fernández-Muñoz, C.A. Martínez, F.A. Pérez and A. Albacete. 2016. Improving agronomic water use efficiency in tomato by rootstock-mediated hormonal regulation of leaf biomass. Plant Sci.251: 90–100.
Costa, J.M. and H. Chall. 2002. The Effect of the Original Leaf Area on Growth of softwood cuttings and planting material of rose. Sci. Hortic. 95(1-2): 111-121.
da Costa, C.T., M.R. de Almeida, C.M. Ruedell, J. Schwambach, F.S. Maraschin and A. G. Fett-Neto. 2013. When stress and development go hand in hand: main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci.133: 1-19.
Druege, U., S. Zerche and R. Kadner. 2004. Nitrogen and storage affected carbohydrate partitioning in high-light-adapted Pelargonium cuttings in relation to survival and adventitious root formation under low light. Ann. Bot. 94(4): 831– 842.
Elhaak, M.A., M.Z. Matter, M.A. Zayed and D.A. Gad. 2015. Propagation Principles in Using Indole-3-Butyric Acid for Rooting Rosemary stem cuttings. J. Hort. 2(1): 121-133.
Esmaeili, M., R. Salehi, M.R. Taheri, M. Babalar and H. Mohammadi. 2015. Effect of different nitrogen rates on fruit yield and quality of grafted and nongrafted muskmelon, in Proceeding of the First International Symposium on Vegetable Grafting. Acta Hortic. 1086: 255–260.
Ezekiel, A. 2010. Viable options and factors in consideration for low cost vegetative propagation of tropical trees. IJB. 6(2): 187-193.
Gudeva, L.K., F. Trajkova, L. Mihajlov and J. Troick. 2017. Influence of different auxins on rooting of rosemary, sage and elderberry. ARRB 12(5): 1-8.
Hansen, J. and K. Kristensen. 2006. Axillary bud growth in relation to adventitious root formation in cuttings. Physiol. Plant. 79(1): 39- 44.
Hartmann, H.T., D.E. Kester, F.T.J. Davies and R.L. Geneve. 2011. Hartmann and Kester’s Plant Propagation: Principles and Practices (8th Ed.). New Jersey: Prentice Hall. p. 928.
Husen, A. 2012. Changes of soluble sugars and enzymatic activities during adventitious rooting in cuttings of Grewia optiva as affected by age of donor plants and auxin treatment. Am. J. Plant Physiol. 7(1): 1-16.
Kerner, N.S., D. Mills and J.C. Merchuk. 2000. Sugar utilization and invertase activity in hairy-root and cell- suspension cultures of Symphytum officinale. Plant Cell Tiss. Org. 62(2): 89-94.
Kochhar, V.K., S.P. Singh, R.S. Katiyar and P. Pushpangadan. 2005. Differential rooting and sprouting behaviour of two Jatropha species and associated physiological and biochemical changes. Curr. Sci. 89(6): 936–939.
Li, H., Y. Wang, Z. Wang, X. Guo, F. Wang, X.J. Xia, J. Zhou, K. Shi, J.Q. Yu and Y.H. Zhou. 2016. Microarray and analysis reveals that csa-miR159b plays a critical role in abscisic acid-mediated heat tolerance in grafted cucumber plants. Plant Cell Environ. 39(8): 1790–1804.
Mason, M.G., J. J. Ross, B. A. Babst, B. N. Wienclaw and C. A. Beveridge. 2014. Sugar demand, not auxin, is the initial regulator of apical dominance. Proc. Nat. Acad. Sci. 111(16): 6092–6097.
Mbangcolo, M.M., E.Y. Reinten and G.A. Agenbag. 2013. Effect of species, cutting position and exogenous rooting substances on rooting of honeybush (Cyclopia spp.) cuttings. South Afr. J. Plant Soil. 30(1): 53-55.
Müller, D., T. Waldie, K. Miyawaki, J. P.C. To, C.W. Melnyk, J.J. Kieber, T. Kakimoto and O. Leyser. 2015. Cytokinin is required for escape but not release from auxin mediated apical dominance. Plant J. 82: 874–886.
Nordström, A., P. Tarkowski, D. Tarkowska, R. Norbaek, C. Åstot, K. Dolezal and G. Sandberg. 2004. Auxin regulation of cytokinin biosynthesis in Arabidopsis thaliana: a factor of potential importance for auxin-cytokinin-regulated development. Proc. Natl. Acad. Sci. 101: 8039– 8044.
Osterc, G., M. Stefancic and F. Stampar. 2009. Juvenile stock plant material enhances root development through higher endogenous auxin level. Acta. Physiol. Plant. 31(5): 899–903.
Osterc, G., M.M. Petkovsek and F. Stampar. 2016. Quantification of IAA metabolites in the early stages of adventitious rooting might be predictive for subsequent differences in rooting responses. Plant Growth Regul. 35(2): 534-542.
Otiende, A. M., J.O. Nyabundi and K. Ngamau.2016. Rose rootstocks position and auxins affect grafting take of ‘Inca’.JAH.18: 54-60.
Otiende, A. M., J.O. Nyabundi, K. Ngamau and P. Opala. 2017. Effects of cutting position of rose rootstock cultivars on rooting and its relationship with mineral nutrient content and endogenous carbohydrates. Sci. Hortic. 225: 204–212
Otiende, A.M., J.O. Nyabundi and K. Ngamau. 2015. Effect of cutting position, auxins and rootstocks on flower yield of rose cultivar ‘Inca’. Acad. Res. Int. 6(4): 9–16.
Pacurar, D. I., I. Perrone and C. Bellini. 2014. Auxin is a central player in the hormone cross-talks that control adventitious rooting. Physiol. Plant. 151(1): 83-96.
Rakibuzzaman, M., K. Shimasaki and A.F.M. JamalUddin. 2018. Influence of cutting position and rooting hormones on rooting of Stevia (Stevia rebaudiana) stem cutting. Int. J. Bus. Soc. Sci. Res. 6(4): 122-125.
Rapaka, V.K., B. Bessler, M. Schreiner and U. Druege. 2005. Inter- play between initial carbohydrate availability, current photosynthesis, and adventitious root formation in Pelargonium cuttings. Plant Sci. 168(6): 1547-1560.
Rasmussen, A., S.A. Hosseini, M.R. Hajirezaei, U. Druege and D. Geelen. 2015. Adventitious rooting declines with the vegetative to reproductive switch and involves a changed auxin homeostasis. Exp. Bot. 66(5): 1437-1452.
Ruedell, C.M., M.R.D. Almeida, J. Schwambach, C. Posenato and A.G. Fett-Neto. 2013. Pre and postseverance effects of light quality on carbohydrate dynamics and Micro adventitious rooting of two Eucalyptus species of contrasting recalcitrance. Plant Growth Regul. 69(3): 235–245.
Rybak-Chmielewska, H. 2007. High performance liquid chromatography (HPLC) study of sugar composition in some kinds of natural honey and winter stores processed by bees from starch syrup. J. Apicult. Sci. 51(1): 23-38.
Saifuddin, M., N. Osman and M.M. Rahman. 2013. Influence of different cutting positions and rooting hormones on root initiation and root-soil matrix of two tree species. Int. J. Agric. Biol. 15: 427-434.
Sevic, H and K. Guney. 2013. Effects of IAA, IBA, NAA and GA3 on rooting and morphological features of Melissa officinalis L. stem cuttings. Sci. World J.1-5.
Shan, X., J. Yan and D. Xie. 2012. Comparison of phytohormone signaling mechanisms. Curr. Opin. Plant Biol. 15(1): 84–91.
Stokes, A., C. Atger, A. Bengough, T. Fourcaud and R. Sidle. 2009. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil. 324(1-2): 1–30.
Suchoff, D., C. Gunter, J. Schulthesis and F.J. Louws. 2015. On farm grafted tomato trial to manage bacterial wilt, in proceeding of the First International Symposium on Vegetable Grafting. Acta. Hortic.1086:119–127.
Thomas, P. and J.W. Schiefelbein. 2004. Roles of leaf in regulation of root and shoot growth from single node softwood cuttings of grape (Vitis vinifera). Ann. Appl. Biol. 144(1): 27-37.
Van Labeke, M.C., P. Dambre, M. Bodson and H. Pien. 2001. Developmental changes in carbohydrate content in young rose shoots (Rosa hybrida ‘Frisco). Acta Hortic. 547: 193–201.
Wendling, I., P.R. Brooks and S.J. Trueman. 2015. Topophysis in Corymbia torelliana × Citriodor seedlings: Adventitious rooting capacity, stem anatomy, and auxin and abscisic acid concentrations. New Forest.46: 107-120.
Wroblewska, K. 2013. Benzyladenine effect on rooting and axillary shoot outgrowth of Gauralin deheimeri Engelm. A. Gray cuttings. Acta. Sci. Pol. Horto. Cultus 12(3): 127-136.
Xu, J., M. Zha, Y. Li, Y. Ding, L. Chen, C. Ding and S. Wang. 2015. The interaction between nitrogen availability and auxin, cytokinin, and strigolactone in the control of shoot branching in rice (Oryza sativa L.). Plant Cell Rep. 34: 1647–1662.

Copyright © Otiende and Maimba, 2020. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium provided the original author and source are appropriately cited and credited.
Join Journal of Environmental and Agricultural Sciences (JEAS)
Interested to join the JEAS Team
Join JEAS as a member Editorial Board see Editors’ Responsibilities
Join JEAS as a member Review Panel Reviewers’ Responsibilities
(send your CV through email at editor.jeas@outlook.com)
JEAS Indexing Journal of Environmental EAS is indexed by reputed indexing services.
Suggest Indexing service/s through email (editor.jeas@outlook.com)
Call for Articles
Submit Your research for publication in the “Journal of Environmental and Agricultural Sciences (JEAS)” through email: editor.jeas@outlook.com
- How to prepare your manuscript before submission
- How to submit your manuscript
- Publication Ethics
- Publication Fee Currently JEAS is publishing manuscripts without publication or processing fee
JEAS Recently Published and Highly Cited Articles
Citation record of JEAS: JEAS Google Scholar page
Follow JEAS Facebook
Thanks for the crucial tips I’ve learned through your blog.